Beispiele für fossil nicht überlieferte Lebensräume und Lazarus-Taxa
Eine Erklärungsmöglichkeit für das Fehlen von Fossilien vieler heutiger Grundtypen in bestimmten Abschnitten der Fossilüberlieferung bietet die Annahme fossil nicht überlieferter Lebensräume. Diese Erklärung besagt, dass manche Taxa (Gruppen von Lebewesen) in Lebensräumen lebten, die (aus verschiedenen Gründen) fossil nicht dokumentiert worden sind. In diesen Lebensräumen hätten heute bekannte Grundtypen (Schöpfungsarten) bereits vor ihrem ersten fossilen Auftauchen schon existiert, wurden aber nicht fossilisiert (vgl. Stephan 2002, 69–71, 80–106; Stephan 2015, 129–131).
Stephan (2002) berichtet ausführlich über die Forschungsgeschichte, die Befunde und auch mögliche Ursachen fossil nicht überlieferter Lebensräume. Übrigens geht diese Theorie auf Georges Cuvier (1769–1832) zurück.
Beispielhafte Befunde von Lücken in der Fossilüberlieferung sind folgende:
Das massive Verschwinden der Quastenflosser (Fleischflosser aus der Gruppe der Actinistia) von vor ca. 70 MrJ (Millionen radiometrische Jahre) bis zu ihrer Wiederentdeckung in den1930er-Jahre gehört mittlerweile sogar zum Allgemeinwissen (Romano et al. 2016, 113; vgl. Stephan 2002, 114–118). Damit sind Quastenflosser nicht nur ein Beispiel für Lebewesen, die lange Zeit in fossil nicht überlieferten Lebensräumen überdauerten, sondern sie sind auch erstaunliche „Lebende Fossilien“: Die heutige Familie der Kahlhechte (Amiidae) hat ein fossiles Alter von 201 MrJ und die heutige Familie der Quastenflosser (Latimeriidae) ist fossil mindestens 241 MrJ alt. Die Ordnung der Coelacanthiformes (Quastenflosserartige) ist mit 388–383 MrJ noch älter (Scholl 2024).

Eine 130 MrJ lange fossile Überlieferungslücke klafft zwischen Odoraia und Wingertshellicus – einer Gruppe „einzigartiger“ Gliederfüßer (Stephan 2002, 69–71).

Ungefähr 50 % der Gattungen, die an der Perm-Trias-Grenze verschwanden, tauchen ca. 9–10 MrJ später wieder auf (S. 74). Nach Massenaussterben der unteren Oberkreide (Cenoman-Turon-Ereignis) traten 50 % der vermeintlich ausgestorbenen Tiere doch später wieder auf (Stephan 2002, 80). Solche in der Fossilüberlieferung zeitweise verschwundenen Arten werden Lazarus-Taxa genannt.
Der Schwamm Ammonella quadrata hat bezüglich des Skelettbaus enge Beziehungen zu Protospongiidae aus dem Erdaltertum, was eine Überlieferungslücke von 210 MrJ (Oberdevon bis Oberjura) vermuten lässt. Auch ein Hornschwamm des Oberjura in Süddeutschland (Nusplinger Plattenkalk) schließt an Formen aus dem Kambrium an – was für eine Lücke von ca. 330 MrJ spricht (Stephan 2002, 84).
Der langarmige Breitschildkrebs Coleia longipes trat im Nusplinger Plattenkalk (ca. 152 MrJ) auf und war zuvor nur aus dem im Toaricum bekannt (ca. 30 MrJ vorher) (Stephan 2002, 84f).
Der Seeigel Paracidaris florigemma im Oberjura taucht nach einer Lücke von 7–8 MrJ wieder auf (Stephan 2002, 85).

Die Nautilusartigen verschwanden zweimal für 2,5 bzw. 5 MrJ aus der Fossilüberlieferung (Stephan 2002, 105f).
Man dachte, dass Urmützenschnecken (Monoplacophora) seit dem Devon vor 375 MrJ ausgestorben seien, allerdings wurden Dutzende Arten der Urmützenschnecken-Gruppe Tryblidiida seit dem Jahr 1952 in der Tiefsee bis ca. 6000 m Tiefe lebendig wiederentdeckt (Stephan 2002, 107f; Lindberg 2009, 191–193). Diese Entdeckung hat „unsere Vorstellung von der Evolution der Mollusca [Weichtiere] kräftig umgestaltet“ (Lindberg 2009, 191).

Weitere interessante Beispiele fossil nicht überlieferter Lebensräume sind: Die Oligozän-Ablagerung Landesplattenberg im Senftal enthält über 3.500 Wirbeltiere, aber seltsamerweise keine Wirbellosen, welche doch existiert haben müssen (Stephan 2002, 92).
Die Gruppe der Schleichenlurche (Gymnophiona) sind heute mit 160 Arten bekannt und weit verbreitet, aber fossil „fast Fehlanzeige“ (Stephan 2002, 99f). Trotz vermuteter Evolutionsgeschichte von ca. 300 MrJ sind erst ab dem Jahr 1973 wenige ältere Fossilien gefunden worden (ebd.).
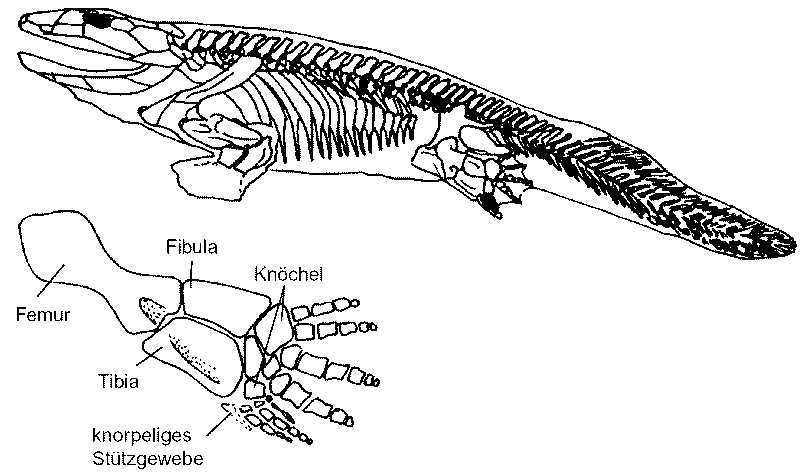
Vor einigen Jahren neu entdeckte Tetrapoden-Spuren waren ca. 18 MrJ älter als die ältesten Körperfossilien der Tetrapoden aus dem frühen Oberdevon wie Acanthostega und Ichthyostega – und ca. 10 MrJ älter als tetrapodenähnliche Fische wie Panderichthys und Tiktaalik aus dem Grenzbereich Mittel-/Oberdevon (Stephan 2015, 127). Obwohl Fußspuren aus Polen (Zachełmie Quarry) mit einem Alter von 390 MrJ aus dem unteren Mitteldevon in Zweifel gezogen wurden, konnten Qvarnström et al. (2018, 1) ihre Identität als früheste fossile Tetrapoden-Spuren mit paläoökologischen Untersuchungen untermauern. Die Autoren sprechen von einem zeitlichen Vorsprung von 14 MrJ zu den ältesten „eindeutigen“ Tetrapoden-Fossilien und von 5 MrJ Vorsprung zu tetrapodenähnlichen Elpistostegalia wie Panderichthys. Laut Barley (2010) in Newscientist sollen die Tetrapodenspuren aus Polen sogar 397 MrJ alt sein.

Ein Kieselschwammriff vor der Westküste Kanadas stellt „ein modernes Analogon“ zu solchen des Jura dar (Stephan 2015, 128f).
Auch bei „praktisch sämtlichen Hauptgruppen der Dinosaurier“ kommen längere Abwesenheitszeiten in der Fossilüberlieferung vor (Stephan 2015, 129f).
Die Chroniosuchier (Reptilomorpha) waren vom Perm bis zur mittleren Trias verschwunden (S. 130).
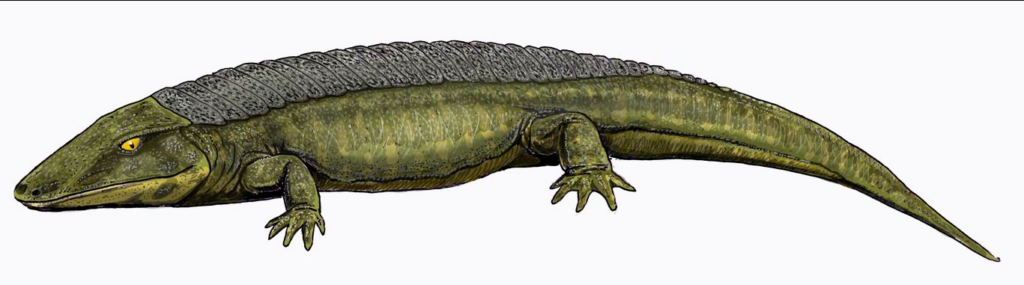
Ähnliches gilt für Echsen der Choristodera-Gruppe, die nicht nur in Jura bis Tertiär (168–11,6 MrJ), sondern auch in der Trias gefunden wurden (Stephan 2015, 130); Schoch (2015, 239–241) weist sogar „eindeutige“ Choristodera in der Mitteltrias Süddeutschlands (Ladinium, 242–237 MrJ) nach, was eine Lücke von 30 MrJ zu Pachystropheus und 80 MrJ zu Cteniogenys in der Fossilüberlieferung bedeutet (ebd.).
Die Nagetierfamilie (Diatomyidae) der in Laos neu entdeckten Laotischen Felsenratte (Leonastes), galt seit dem Miozän (11 MrJ) als ausgestorben (Stephan 2015, 130f).

In Bezug auf die Fische lässt sich beispielhaft ergänzen, dass die Katzenhai-Gattung Thiesus von Bathonium (166 MrJ) bis Valenginium (140 MrJ) aus der Fossilüberlieferung verschwand; sie gehörte also 25 MrJ lang zu den Lazarus-Taxa (Guinot et al. 2014, Fig. 12).
Für Elasmobranchii (Plattenkiemer: Haie und Rochen) scheint die Fossilüberlieferung übrigens in Bezug auf die Daten heutiger Taxa von Paillard et al. (2020, 4) ziemlich vollständig zu sein: 100 % der Ordnungen, 88 % der Familien und 56 % der Gattungen sind fossil vertreten. Diese fossil vertretenen heutigen Gattungen existieren im Mittelwert schon seit ca. 57 MrJ (vgl. ebd., supinfo1).
Zahlreiche weitere Beispiele in Bezug auf Insekten aus der Kreide finden sich bei Suling (2021): „Viele der heute noch lebenden Familien, die man vor dem Albium [ = Stufe der Unterkreide] […] allein aus Sedimenteinschlüssen kennt, sind in der Oberkreide gar nicht oder allein als Bernsteineinschlüsse nachgewiesen.“ (ebd., 3).
Schlussfolgerungen
Lücken in der Fossilüberlieferung sind also keine Einzelfälle, sondern ein typischer, regelmäßiger Befund der fossilen Überlieferung, mit dem sich sowohl Forscher von Evolutions- als auch Grundtypmodellen auseinandersetzen müssen – wenn auch aus jeweils anderen Gründen (s. Scholl 2024). Das Auftreten von fossil früh bekannten und dann später zeitweise in der Fossilüberlieferung wieder verschwundenen Lazarus-Taxa betrifft beide Perspektiven gleichermaßen.
Für evolutionäre Modelle existiert insgesamt – so wie z. B. bei Fischgruppen des Silur (Scholl 2024) – das systematische Problem, dass unzählige zu erwartende Übergangsformen zu den richtigen Zeitpunkten in der Fossilüberlieferung fehlen. Neu entdeckte, noch ältere Fossilfunde, die wiederum häufig unbekannte und unerwartete Merkmalsmosaike aufweisen, lösen das Problem von zu überbrückenden Bauplanunterschiede in der Regel nicht, sondern verlagern es zeitlich nur nach hinten (und vergrößern es in manchen Fällen), da hier wieder von anderen bisher unbekannten Übergangsformen ausgegangen werden muss. Dies wird auch darin deutlich, dass Ausdrücke wie „unerwarteter Merkmalsmix“ oder „die Evolutionsgeschichte der Gruppe XY muss neu geschrieben werden“ nahezu standardmäßig in Meldungen über neue Fossilfunde vorkommen (Scholl 2024).
Schöpfungsmodelle hingegen müssen sich mit der folgenden Frage auseinandersetzen: Warum treten moderne Großgruppen zwar typischerweise fossil früh in Erscheinung, während viele Gruppen mittlerer systematischer Ebenen (Ordnungen und v. a. Familien), die wahrscheinlich am ehesten Schöpfungseinheiten bzw. Grundtypen entsprechen (vgl. Hartmann & Junker 2009; ähnlich bei Behe 2019, 152–156), jedoch häufig fossil relativ jung sind? Vgl. zu dieser Thematik ausführlich Scholl 2024, Stephan 2002 und 2015. Ausnahmen sind allerdings zahlreiche Beispiele lebender Fossilien (wie z. B. Neunaugen seit 360 MrJ), die zumindest in ihrem äußerlichen Körperbauplan der Evolution für Dutzende oder Hunderte Millionen Jahre im Wesentlichen getrotzt haben.
Literatur
Barley S (2010) Oldest footprints of a four-legged vertebrate discovered, vom 06.01.2010, https://www.newscientist.com/article/dn18346-oldest-footprints-of-a-four-legged-vertebrate-discovered/.
Behe M (2019) Darwin Devolves: The New Science About DNA That Challenges Evolution (English Edition). Kindle Ausgabe. HarperOne.
Guinot G, Capetta H & Adnet S (2014) A rare elasmobranch assemblage from the Valanginian (Lower Cretaceous) of southern France. Cretaceous Research 48, 54–84, doi: 10.1016/j.cretres.2013.11.014.
Hartmann F & Junker R (2009) Paßten alle Tiere in die Arche Noah? W+W Diskussionsbeitrag 4/90, letzte Aktualisierung 10.09.2009
Lindberg DR (2009) Monoplacophorans and the origin and relationships of mollusks. Evo. Edu. Outreach 2, 191–203, doi: 10.1007/s12052-009-0125-4, https://escholarship.org/content/qt342063m8/ qt342063m8. pdf.
Qvarnström M et al. (2018) Non-marine palaeoenvironment associated to the earliest tetrapod tracks. Sci. Rep. 8, 1074, https://doi.org/10.1038/s41598-018-19220-5.
Schoch RR (2015) Reptilien des Lettenkeupers. In: Hagdorn H, Schoch R & Schweigert G (Hrsg.): Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern. Staatliches Museum für Naturkunde Stuttgart, 231–264, http://www.palaeodiversity.org/pdf/08Suppl/10Palaeodiversity_SB_Schoch.pdf.
Scholl B (2024) Die ältesten fossilen Fische: Große Vielfalt seit Beginn der fossilen Überlieferung. W+W-Onlineartikel, https://www.wort-und-wissen.org/artikel/die-aeltesten-fossilen-fische/.
Stephan M (2002) Der Mensch und die geologische Zeittafel. Warum kommen Menschenfossilien nur in den obersten geologischen Schichten vor? SCM Hänssler.
Stephan M (Hrsg.) (2015) Sintflut und Geologie. Schritte zu einer biblisch-urgeschichtlichen Geologie. 4. Aufl. SCM Hänssler.
Suling M (2021) Kretazische Insekten im Gestein und als Einschlüsse. W+W Special Paper B-21-2, https://www.wort-und-wissen.org/wp-content/uploads/b-21-2_kretazische-insekten.pdf.
Dieser Artikel ist ein leicht überarbeiteter Auszug aus dem Online-Artikel: Scholl B (2024) Die ältesten fossilen Fische: Große Vielfalt seit Beginn der fossilen Überlieferung. W+W-Onlineartikel, https://www.wort-und-wissen.org/artikel/die-aeltesten-fossilen-fische/.