Rudimentäre Organe
Viele Organe und Strukturen der Lebewesen scheinen nicht optimal konstruiert zu sein. Es wird in diesem Fällen vermutet, dass sie im Laufe der Evolution zurückgebildet (rudimentiert) seien oder dass aufgrund des mutmaßlichen evolutionären Umbaus keine optimalen Lösungen möglich waren. Diese Argumentation lässt sich jedoch nicht stichhaltig begründen.
1.0 Inhalt
In diesem Artikel wird das evolutionstheoretische Argumentation der Rudimentation vorgestellt und Kriterien zur Erkennung von rudimentären Organen diskutiert. Außerdem wird auf die Behauptung eingegangen, es gäbe Konstruktionsfehler im Bau der Lebewesen.
1.1 Einleitung
Wenn die Lebewesen evolutionär entstehen, müssen nicht nur Organe ganz neu gebildet werden, sondern es ist zu erwarten, dass vorhandene Organe umgebaut oder auch im Laufe der Zeit wieder abgebaut werden. Ein evolutionärer Prozess kann ja nicht „wegen Umbau schließen“ (G. Osche), vielmehr müssen die Lebewesen in jeder Generation überlebensfähig sein. Es wird im hypothetischen Prozess der Evolution – bildhaft gesprochen – bevorzugt an dem „weitergebaut“, was schon da ist: Umbau statt Neubau. Oder: Wenn eine Struktur z. B. aufgrund geänderter Umweltbedingungen oder nach Eroberung eines neuen Lebensraums nicht mehr benötigt wird, so kann sie im Laufe der Zeit verkümmern oder ganz verschwinden.
An diesen Gedankengängen knüpfen zwei evolutionäre Konzepte an: das der Rudimentation und das der „Konstruktionsfehler“.
1.2 Rudimentation
Als „rudimentär“ werden in der Evolutionsbiologie Organe bezeichnet, die keine Funktion oder eine im Vergleich zu ihrem Bau zu geringfügige Funktion besitzen sollen. Solche Organe scheinen eine Unstimmigkeit zwischen Struktur und Funktion aufzuweisen: Der strukturelle Aufwand scheint für die zu erfüllende Funktion zu hoch zu sein. Berühmtes Beispiel beim Menschen ist der Wurmfortsatz (Appendix) des Blinddarms (Abb. 73).
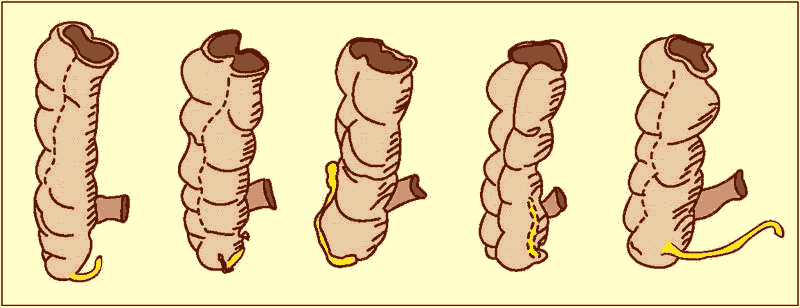
Abb. 73: Der Wurmfortsatz am Blinddarm des Menschen mit fünf verschiedenen Größen und Lagen. Quelle: Nach KRUMBIEGEL.
Rudimentation kann man folglich als „Rückbildung“ bezeichnen. Ein Organ wird als „rudimentär“ interpretiert, wenn es durch Rückbildung eines ehemals funktions-
tüchtigeren Organs entstanden sein soll. Als Grund für eine Rudimentation wird gewöhnlich der Wegfall von Selektion angesehen. Wenn aufgrund der Änderung der Lebensweise ein Organ nicht mehr benötigt wird, werden schädliche Mutationen nicht mehr ausgelesen; sie sammeln sich an und führen zur Verkümmerung des betreffenden Organs. In diesem Sinne gelten rudimentäre Organe als Belege für eine regressive Evolution. (Es ist zu beachten, dass der Vorgang der Rudimentation in der Regel nicht beobachtet wurde, sondern anhand von Indizien erschlossen wird, dazu weiter unten im Abschnitt „Woran erkennt man rudimentäre Organe?“ mehr.)
Der eben beschriebene Vorgang ist an sich natürlich kein Beleg für eine konstruktive Evolution (s. „Mikro- und Makroevolution“ (https://genesis-net.de/e/1-3-a/1-3/)), denn durch Rudimentation kann allenfalls der Verlust eines Organs erklärt werden. Rudimentation, für sich alleine betrachtet, ist degenerative Mikroevolution.
Der Hinweis, Rudimentation passe nicht zu einer Schöpfungsvorstellung (s. Abschnitt „Theologische Argumentation“), kann nicht überzeugen, denn ein Schöpfungsmodell schließt Rudimentation nicht generell aus (s.u.).
Viele Beispiele von rudimentären Organen können als bloße Degenerationen gedeutet werden, etwa die Verkümmerung von Augen bei höhlenlebenden Fischen (Abb. 74) oder die Verkümmerung von Insektenflügeln in speziellen Standorten.

Abb. 74: Flussfisch Astyanax mexicanus und die blinde Höhlenform. Beide gehören zur selben Biospezies.
Kopplung von Rudimentation mit Neukonstruktion. Die Situation ist allerdings oft komplizierter, wenn das rückgebildete Organe nicht isoliert betrachtet wird. Das Beispiel der blinden Höhlensalmler (Abb. 74) kann die Problematik verdeutlichen. Diese Fische leben in stockfinsteren Höhlen. Das Augenlicht würde ihnen dort nichts nützen. Dennoch müssen sie sich orientieren können. Dazu dient ihnen u. a. ein Geschmacksknospenfeld, das um das Maul herum ausgebildet ist. Solche Geschmacksknospen besitzen alle Salmler.
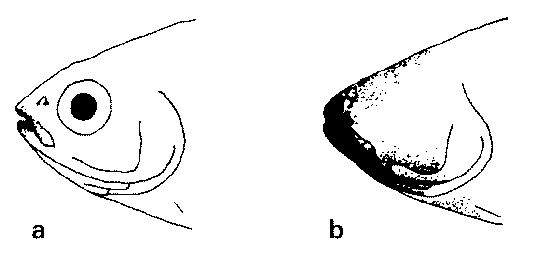
Abb. 75: Geschmacksknospenbesatz (a) von Astyanax und (b) von der blinden Höhlenform. Nach PETERS.
Die höhlenlebenden Formen haben aber ein deutlich vergrößertes Geschmacksknospenfeld (Abb. 75). Dadurch wird die verlorene Orientierung kompensiert. Genetische Studien zeigten, dass die Vergrößerung des Geschmacksknospenfeldes dadurch erfolgte, dass Modifikatorgene (durch die die Ausbildung von Geschmacksknospen unterdrückt wird) durch Mutation außer Kraft gesetzt wurden. Dadurch wird das genetische Programm für die Ausbildung von Geschmacksknospen auf der Kopfoberfläche in größerem Ausmaß abgerufen. Die Erweiterung des Geschmacksknospenfeldes beruht also nicht auf einer konstruktiven Änderung, sondern auf einem Ausfall in der Steuerung. Neue Strukturen oder neue Gene sind nicht gebildet worden.
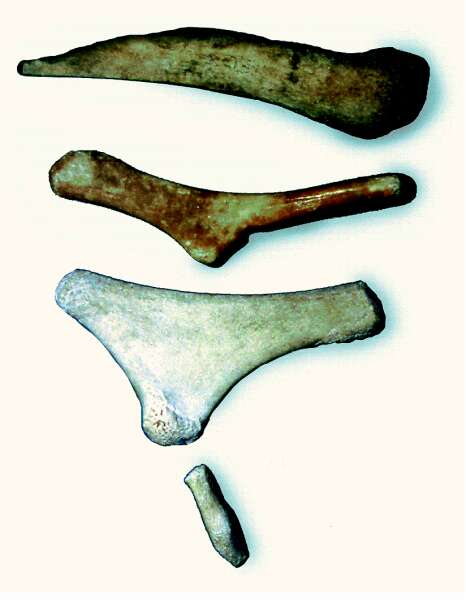
Abb. 76: Abdominalknochen des Pottwals, des Seiwals und des Finnwals (von oben nach unten). Der Finnwal besitzt außerdem einen als Oberschenkelrudiment gedeuteten Knochen.
Die Bauchknochen der Wale: Kompensation durch Makroevolution? Nicht alle Kompensationen können – wie im obigen Beispiel – durch mikroevolutive Prozesse erklärt werden. Die mit der Rückbildung gekoppelte Neubildung anderer Organe würde – wenn sie auf evolutiven Wege erfolgt ist – oftmals makroevolutive Änderungen erfordern. Ein berühmtes Beispiel sind die Bauchknochen (sog. Abdominalknochen) der Walartigen. Es handelt sich um paarige Knochen, die in der Muskelmasse des Walbauches stecken, und zwar etwa dort, wo sich bei landlebenden Säugetieren das Becken befindet (Abb. 76) – ohne Verbindung zur Wirbelsäule. Manche Walartige besitzen ein weiteres Knochenpaar; der Grönlandwal sogar ein drittes (Abb. 77). Evolutionär werden die Knochen als Rudimente eines ehemaligen Beckens und ggf. als Reste der Hinterbeine interpretiert. Somit werden diese Knochen als Hinweise für einen Übergang von Landsäugetieren zu den Walen gewertet. Bei einem solchen Übergang müsste aber ein Großteil des Körpers tiefgreifend umgebaut worden sein – es würde sich dabei zweifellos um Makroevolution handeln. Also: Rudimentation (Becken, Hinterbeine) und Makroevolution (Umkonstruktion; viele Organe) müssten irgendwie Hand in Hand gegangen sein. In solchen Fällen werden rudimentäre Organe als indirekte Belege für eine Makroevolution gewertet.
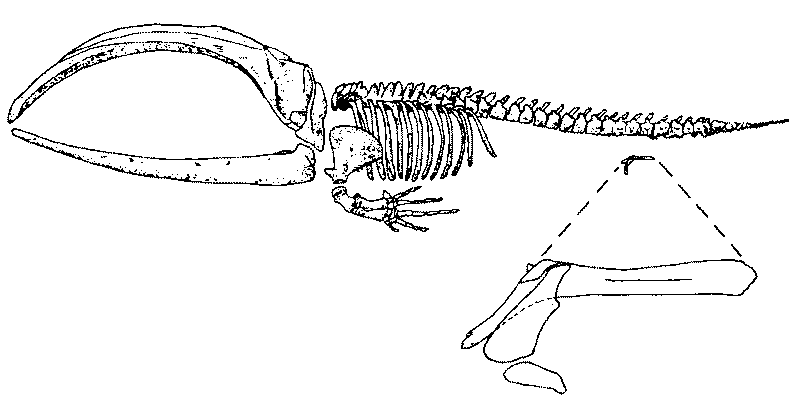
Abb. 77: Skelett des Grönlandwals (Balaena mysticetus) mit Becken- und Hinterextremitätenrudiment. Nach MÜLLER.
Allerdings kann man hier kritisch zurückfragen, ob die Rudimentation überhaupt empirisch belegt ist. Woran erkennt man, dass die Bauchknochen der Wale zurückgebildete Becken- (und Bein-)knochen sind? Damit kommen wir zum nächsten Punkt.
1.3 Woran erkennt man rudimentäre Organe?
Wenn eine Art, das ein Organ in voll ausgebildeter Form besitzt, mit einer anderen Art, die dieses Organ nur rudimentär besitzt, kreuzbar ist, so kann die Kreuzbarkeit als Indiz für eine tatsächlich erfolgte Rückbildung gelten. Dies ist beispielsweise der Fall beim blinden Höhlensalmler, der mit den sehtüchtigen Formen problemlos kreuzbar ist (vgl. Abb. 78).

Abb. 78: Auswahl aus der 2. Generation einer Kreuzung des blinden Höhlensalmlers und einer sehtüchtigen Form, mit Übergängen von blind bis sehend. Zeitschrift für zoologische Systematik und Evolutionsforschung, Band 1, 1963.
Als weiteres typisches Beispiel seien die Weisheitszähne des Menschen angeführt, die oft ganz spät durchbrechen. Man kann davon ausgehen, dass ursprünglich alle 32 Zähne des Menschen regelmäßig voll funktionstüchtig waren und auch gebraucht wurden. Dass der heutige Mensch die Weisheitszähne nicht mehr unbedingt braucht, hängt möglicherweise mit seinen veränderten Essgewohnheiten zusammen. Daher war eine „degenerative“ Entwicklung dieses Backenzahns nicht nachteilig und deshalb möglich. Die Rückbildung der Weisheitszähne ist übrigens bei verschiedenen Menschen unterschiedlich. Als drittes Beispiel seien flugunfähige Insekten genannt, die auf windumtosten Inseln leben (s. „Mutation“ (https://genesis-net.de/e/1-3-b/2-1/) und Abb. 58).
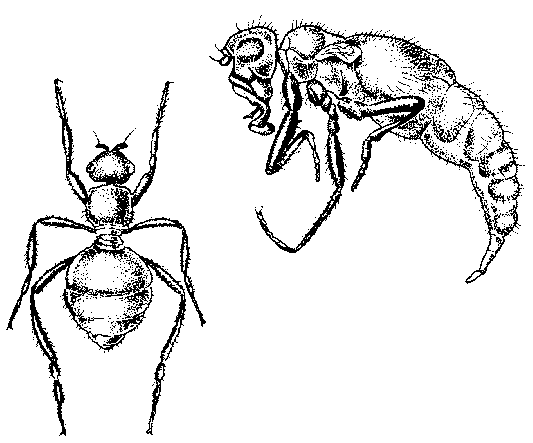
Abb. 58: Zwei Beispiele flugunfähiger Insekten, die auf windumtosten Inseln vorkommen: Tangfliege (links), Dungfliege (rechts). R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
Im Falle der Wale greift das Kriterium der Kreuzbarkeit nicht. Dafür sind sie viel zu verschieden von ihren mutmaßlichen landlebenden Vorfahren entfernt. Wie wird dann aber solchen Fällen eine Rückbildung erkannt? Im Wesentlichen gibt es hier-
zu zwei Kriterien:
1. Funktionslosigkeit. Könnte man nachweisen, dass ein Organ gar keine Funktion hat, also vollkommen nutzlos ist, könnte dies als Hinweis darauf gewertet werden, dass es früher einmal eine Funktion hatte und diese verloren hat. Die Bauchknochen der Wale kann man damit allerdings nicht als rudimentär erkennen, denn sie haben eine sehr wichtige Funktion: Sie bilden Ansatzstellen für die Muskulatur der Geschlechtsorgane und möglicherweise für den Hebermuskel des Afters. Sie unterstützen außerdem die Bauchwand und die Eingeweide. Im Übrigen ist Funktionslosigkeit schwer nachweisbar. Man kann oft nur sagen, dass eine Funktion unbekannt ist, nicht aber, dass es gar keine gibt. Schon in vielen Fällen wurden lebenswichtige oder mindestens nützliche Funktionen von Organen erkannt, die zuvor als funktionslos galten. Das Argument der Funktionslosigkeit ist also problematisch und nicht stichhaltig.
Eine Studie der amerikanischen Forscher William Parker und Mitarbeiter brachte neuerdings bezüglich der Funktion des Wurmfortsatzes des Menschen das Ergebnis, dass der Fortsatz eine Art Zufluchtsort („safe house“) und Rettungsstation für symbiotische Bakterien darstellt, die das Wachstum nützlicher Darmbakterien fördert und bei durchfallbedingten Darmentleerungen die Wiederbesiedlung mit diesen Bakterien ermöglicht bzw. erleichtert. Diesen Bakterien fällt die Aufgabe zu, die Ausbreitung gefährlicher Krankheitserreger im menschlichen Verdauungstrakt zu verhindern, was besonders nach Durchfallerkrankungen sehr wichtig ist. Details werden in „Der Wurmfortsatz als Rettungsstation“, https://genesis-net.de/wp-admin/post.php?post=2616&action=edit erklärt.
Unstimmigkeit von Struktur und Funktion. Eine gegenüber dem Argument der Funktionslosigkeit abgeschwächte Argumentation lautet, dass rudimentäre Organe eine im Vergleich zum strukturellen Aufwand zu geringe Funktion haben. Aber auch eine solche Unstimmigkeit von Struktur und Funktion ist kaum nachweisbar.
2. Formenvergleich mit homologen Organen. In zahlreichen Fällen wird die mutmaßliche Rudimentation durch einen Vergleich mit homologen Organen [= vereinfacht: bauplangleiche Organ] begründet (zum Thema Homologie siehe „Ähnlichkeiten in der Morphologie und Anatomie“ (https://genesis-net.de/e/1-3-e/5-1/)). Zwei Arten werden verglichen, von denen die eine ein homologes Organ in voller Ausprägung, die andere in rudimentärer Form besitzt. Wir können nochmals das instruktive Beispiel der Walartigen heranziehen. Deren Bauchknochen (Abb. 77) üben im Vergleich zu den homologen Skeletteilen der Landwirbeltiere zwar auch Funktionen aus (s.o.), aber diese sind anders ausgerichtet als bei den als homolog betrachteten Beckenknochen von Landsäugetieren.

Abb. 77: Skelett des Grönlandwals (Balaena mysticetus) mit Becken- und Hinterextremitätenrudiment. Nach MÜLLER.
Die evolutionäre Argumentation lautet: Die rudimentären Becken- und Oberschenkelknochen der Wale haben zwar eine spezifische Funktion, aber es liegt eine homologe Ähnlichkeit dieser Strukturen mit den entsprechenden Knochen der Landsäugetiere vor. Damit wird diese Ähnlichkeit als Hinweis auf einen Abstammungszusammenhang gewertet. Das eigentliche Argument für einen Abstammungszusammenhang ist hier also die Beobachtung von Ähnlichkeit, die jedoch, wie im Artikel „Ähnlichkeiten in der Morphologie und Anatomie“ (https://genesis-net.de/e/1-3-e/5-1/) gezeigt, keinen unabhängigen Beleg für Makroevolution darstellt.
Teilweiser Funktionsverlust? Damit ist auch das Argument angesprochen, rudimentäre Organe könnten daran erkannt werden, dass sie gegenüber früheren Ausprägungen noch Restfunktionen besäßen. Die Bauchknochen der Walartigen bieten für dieses Argument nochmals ein geeignetes Anschauungsmaterial. Der in diesem Fall evolutionstheoretisch angenommene Funktionsverlust, ausgehend vom Becken und den Beinen der Landsäugetiere, ist keine Beobachtungstatsache, sondern Deutung unter Vorgabe der Evolutionstheorie. Dass die Bauchknochen der Walartigen in deren evolutionstheoretisch postulierten Vorfahren als Beckengerüst ausgebildet waren, kann an der Ausprägung dieser Skelettelemente nicht ab-gelesen werden. Diese Deutung ist zwar im evolutionstheoretischen Kontext möglich, aber nicht zwingend. Genauso gut kann hier nämlich angenommen werden, dass die Bauchknochen im Wesentlichen in der Ausprägung geschaffen wurden (mit art- und gattungsspezifischen Unterschieden), in der sie heute vorliegen – eine Ausprägung, die für die walartige Lebensweise passend und aufgrund der heutigen Lebensweise verständlich ist.
1.4 Konstruktionsfehler
Manche Strukturen werden als stammesgeschichtlich bedingte Fehlkonstruktionen angesehen (vgl. dazu auch den Artikel „Argumente gegen Design“ (https://genesis-net.de/s/0-4/2-1/)). Es handelt sich in diesen Fällen nicht um Rudimentation im Sinne von Rückbildungen, sondern um Organe, die, wie man glaubt, Spuren eines evolutionären Umbaus zeigen. Es wird argumentiert, dass im Laufe der Evolution oft nur umgebaut werden könne, was zu suboptimalen Konstruktionen und regelrechten Konstruktionsfehlern führe. In diesem Sinne wird ein Argument der Unvollkommenheit eingeführt. Solche Unvollkommenheiten seien Indizien für Evolution, da beim evolutionären Umbau oft nur unvollkommene Lösungen möglich seien.
Als Beispiel hierfür wird gelegentlich die Überkreuzung von Speise- und Luftröhre bei Säugetieren genannt. Wegen der Gefahr des Verschluckens sei diese Konstruktion nicht optimal und als stammesgeschichtlich bedingte Fehlkonstruktion anzusehen. Doch davon kann nicht die Rede sein, denn bei Nicht-Überkreuzung würde die Speiseröhre vor dem Herzen liegen, was z. B. bei einer Vergrößerung des Herzens zu einem Abdrücken der Speiseröhre führen würde. Die Überkreuzung hat zudem einige Vorteile: In der Luftröhre hinaufbeförderter Schleim kann in die Speiseröhre abgeleitet werden. Außerdem macht diese Konstruktion Atmung durch den Mund möglich, was bei Anstrengung, beim Heraushusten von Fremdkörpern oder auch bei starkem Schnupfen eine dankenswerte Einrichtung ist. Außerdem ist diese Konstruktion platzsparend. Eine Unstimmigkeit von Struktur und Funktion liegt nicht vor.
Ein viel zitiertes Beispiel ist in diesem Zusammenhang die Netzhaut im Augenhintergrund der Wirbeltiere. Die lichtempfindliche Zellschicht liegt im Auge sozusagen „falsch“ (invers) herum, so dass das eintretende Licht zuerst die abführenden Nervenbahnen und Zellschichten durchlaufen muss, bevor es auf die lichtempfindlichen Stellen der Netzhaut trifft (Abb. 79, rechts). Anders herum scheint es besser zu sein (Abb. 79, links).
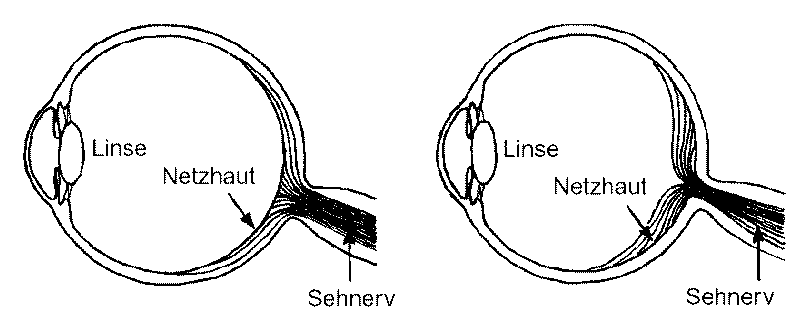
Abb. 79: Das menschliche Auge, wie es gebaut sein sollte, wenn die Netzhaut „richtig“ herum eingelegt wäre (links; direkt dem einfallenden Licht zugewandt), und wie es tatsächlich gebaut ist (rechts). Nach NESSE & WILLIAMS.
Tatsächlich aber erfüllt das Auge seine Aufgaben trotz dieses „Fehlers“ hervorragend. Die inverse Lage hat den Vorteil, dass Streulicht ausgefiltert wird. Detaillierte Studien der Netzhaut und ihrer Funktionsweise haben keine Konstruktionsfehler erweisen können. Der eigentliche Fehler liegt hier in der isolierten Betrachtungsweise einer Detailstruktur unter Ausblendung des ganzen Organs bzw. des ganzen Organismus und auch seiner Ontogenese [= individuelle Entwicklung]. Mit dem Argument der Unvollkommenheit wird häufig vorschnell argumentiert, noch bevor genauere Funktionszusammenhänge erschlossen sind.
Im Übrigen muss bedacht werden, dass man kaum sagen kann, ob es überhaupt eine Alternative gibt, die besser funktioniert, solange keine bessere Alternative erprobt ist.
Details dazu werden in diesem Artikel erläutert: Zankapfel Auge. Ein Paradebeispiel für „Intelligent Design“ in der Kritik (http://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-1.html)
1.5 Theologische Argumentation
Die Argumentation mit „Rudimentation“ und „Konstruktionsfehlern“ wird häufig mit einer Kritik an Schöpfungsvorstellungen verknüpft. So wird nicht selten behauptet, rudimentäre Organe sprächen gegen eine durchdachte Schöpfung, da ein Schöpfer keine nutzlosen Organe erschaffen würde. Ähnlich wird mit dem Argument der Unvollkommenheit (s. o.) verfahren. Daraus wird dann ein indirektes Argument für Evolution gemacht (was gegen Schöpfung spricht, stützt eine evolutionäre Weltsicht). Diese Argumentation ist theologischer Natur, denn sie beruft sich auf Vorstellungen über Handlungsweisen eines Schöpfers: Ein Schöpfer würde keine nutzlosen Organe erschaffen. Eine theologische Argumentation hat aber im Rahmen von Evolutionstheorien keinen Platz. Im Rahmen von Schöpfungsvorstellungen hat diese Frage natürlich ihr Recht (vgl. „Argumente gegen Design“ (https://genesis-net.de/s/0-4/2-1/) und „Todesstrukturen in der Schöpfung“ (https://genesis-net.de/s/0-5/2-1/)).
Es ist zudem zu bedenken, dass Argumente gegen Schöpfung nicht automatisch Argumente für Evolution sind. Und schließlich ist – wie erwähnt – der Nachweis einer Funktionslosigkeit oder Unvollkommenheit schwierig; entsprechende Behauptungen wurden schon oft widerlegt.
1.6 Zusammenfassung
Organe können unter bestimmten Umweltbedingungen rudimentieren und da-
durch ihre Funktion mikroevolutiv teilweise oder ganz einbüßen.
Viele sog. „rudimentäre Organe“ erfüllen eine Funktion, die oft erst durch genauere Untersuchungen aufgedeckt werden konnte. Ähnlichkeiten dieser Organe mit solchen mutmaßlichen stammesgeschichtlichen Vorfahren stellen keinen unabhängigen Beleg für Makroevolution dar.
Funktionslosigkeit von Organen und Konstruktionsfehler sind kaum nachweisbar und in vielen Fällen zu Unrecht behauptet worden.
Autor: Reinhard Junker, 18.03.2008
Aktualisiert am 07.01.2024 (B. Scholl); © beim Autor; alter Link: 2008, https://www.genesisnet.info/schoepfung_evolution/i41303.php
Aktuelle Wissenschaftsnachrichten zu dem Themenbereich lesen.
Hier geht es zu häufig gestellten Fragen zu diesem Thema.
Zurück zu „Vergleichende Biologie“.