Ähnlichkeiten in Morphologie & Anatomie (Experten)
2.0 Inhalt
Die Ähnlichkeiten zwischen den Lebewesen sind für die Evolutionslehre von zentraler Bedeutung. Zum einen gelten sie als wesentliche Belege für Makroevolution. Zum anderen bilden sie die wichtigste Grundlage für Stammbaumrekonstruktionen. Im Folgenden wird gezeigt, dass der „Ähnlichkeitsbeweis“ für Evolution nicht stichhaltig ist und dass eine Reihe von Ähnlichkeitsbefunden evolutionstheoretisch schwer deutbar sind.
2.1 Einleitung
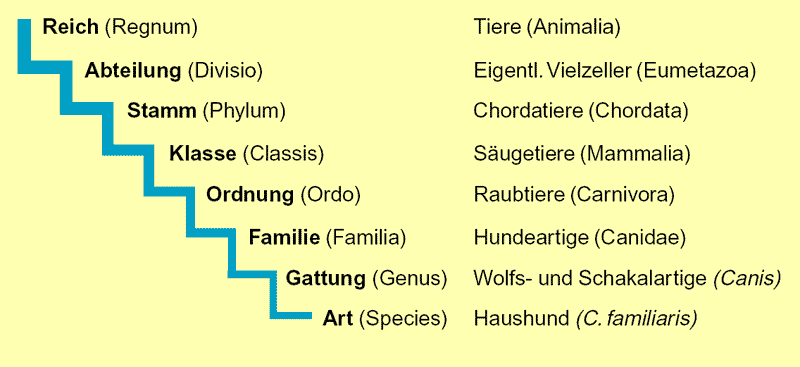
Abb. 1: Die verschiedenen hierarchischen Klassifikationsebenen am Beispiel der Hundeartigen.
Die Ähnlichkeiten der Lebewesen gelten als besonders wichtige Belege für die Abstammung der Arten und für eine allgemeine Evolution (Abb. 1: Klassifikationsebene der Hundeartigen und Abb. 2: Vergleich von Vordergliedmaßen). Folglich gehört der „Ähnlichkeitsbeweis“ zu den wichtigsten Stützen der Evolutionslehre.
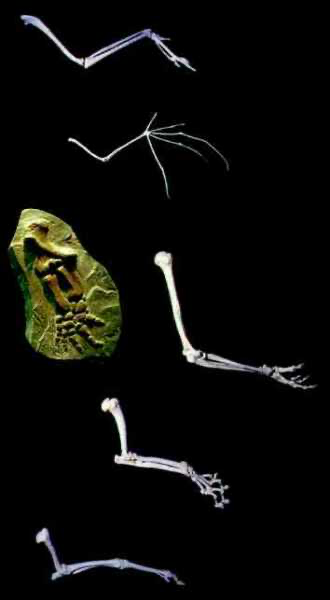
Abb. 2: Das klassische Beispiel für Bauplanähnlichkeiten, die gewöhnlich auf gemeinsame Abstammung zurückgeführt werden. Quelle: Nach Objekten aus dem Westfälischen Museum für Naturkunde, Münster.
Das zugrundeliegende Argument ist einfach und eingängig: Aus Erfahrung wissen wir, dass durch Vererbung ähnliche Merkmale weitergegeben werden: Kinder gleichen ihren Eltern, weil sie von ihnen abstammen. Ähnlichkeit folgt also aus Abstammung und Vererbung. Durch Vererbung pflanzen sich Ähnlichkeiten in den Nachkommen fort. Das können wir auch beobachten – allerdings nur innerhalb von Arten. Dort sind Kreuzungen und Nachkommenproduktion möglich und die Aufeinanderfolge der Generationen und die Ähnlichkeiten in der Generationenfolge können direkt untersucht werden.
Das Argument „Ähnlichkeit durch Abstammung“ wird nun auf verschiedene Grundtypen (z. B. Schimpanse und Mensch) ausgeweitet. Deren Abstammungszusammenhang kann aber nicht mehr beobachtet werden. (Es gibt keine beobachtete Kette von affenartigen Vorfahren des Menschen bis zum Menschen (Paläanthropologie). Das Zustandekommen der Ähnlichkeit beispielsweise zwischen Schimpanse und Mensch ist also keine Folgerung aus Beobachtungen. Vielmehr wird eine Schlussfolgerung vom beobachtbaren Bereich (Nachkommenproduktion innerhalb von Arten) auf den nicht-beobachtbaren Bereich (Abfolge von einem Grundtyp zu einem anderen) gezogen.
Dabei handelt es sich um einen sogenannten Analogieschluss, d. h. einer Schlussfolgerung von Bekanntem auf Unbekanntes, z. B. von einem kleinen Bereich auf einen viel größeren. Hier wird von den Verhältnissen innerhalb einer Art bzw. eines Grundtyps auf die Verhältnisse darüber hinaus geschlossen. Eine solche Schlussfolgerung mag plausibel erscheinen, sie ist jedoch grundsätzlich nicht zwingend. Ob Ähnlichkeiten über den experimentell nachvollziehbaren Bereich hinaus auf Abstammung hinweisen, bleibt daher offen. Es handelt sich um eine Deutungsmöglichkeit, nicht aber um einen Deutungszwang.
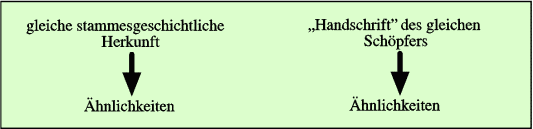
Abb. 3: Zwei mögliche Begründungen für das Vorliegen von Ähnlichkeiten.
Ähnlichkeiten können nämlich auch anders erklärt werden (vgl. Abb. 2):
- aufgrund ähnlicher Funktionen. Beispiel: Die Tiere atmen dieselbe Luft, fressen vergleichbare Nahrung usw., folglich sind ähnliche Organe, Stoffwechseleigenschaften, Erbanlagen (Gene) etc. zu erwarten.
- durch die „Handschrift“ desselben Schöpfers. So wie ein Konstrukteur ähnliche Bauelemente für verschiedene Konstruktionen verwendet, können auch Ähnlichkeitsmuster der Lebewesen als Hinweis auf einen gemeinsamen Urheber interpretiert werden (vgl. Abb. 4). Auch hierbei handelt es sich um einen Analogieschluss, der als solcher auch in diesem Fall nicht zwingend ist.

Abb. 4: Spezielle Ähnlichkeiten von VW-Käfer und Porsche weisen auf dieselben Konstrukteure hin, die beide Modelle gebaut haben.
Gegen diese beiden Erklärungen werden zwei Einwände erhoben:
2.2 Einwand 1: Viele Ähnlichkeiten können nicht durch die Funktion erklärt werden.
Diese nicht-funktionsbedingten Ähnlichkeiten müssen eine andere Ursache haben. Dafür kommt nur gemeinsame Abstammung in Frage. Ein Beispiel: Das Skelett der Brustflosse der Wale und Delphine weist im Bauplan auffallende Ähnlichkeiten mit dem Bauplan des Gliedmaßenskeletts der Landsäugetiere auf. Dagegen haben die Fische einen anderen Bauplan des Flossenskeletts, obwohl dieses dieselbe Funktion ausübt. Die Ähnlichkeiten des Flossenskeletts der Wassersäuger mit dem Skelett der Gliedmaßen der Landwirbeltiere erklärt sich also nicht aus der Funktion, sondern aus der Abstammung der Wassersäuger (Wale, Delphine) von Landsäugetieren.
Erwiderung: Dieses Argument ist nicht stichhaltig: 1. Es kann kaum sicher nachgewiesen werden, dass bestimmte Ähnlichkeiten oder Unterschiede nicht funktionsbedingt sind. Es kann sein, dass ein funktioneller Grund für die in Rede stehende Ähnlichkeit noch nicht gefunden wurde. Angesichts großer Unkenntnisse über die Struktur-Funktions-Kenntnisse beruht eine Aussage wie “Diese Struktur ist durch ihre Funktion nicht (voll) verstehbar“ wahrscheinlich auf Unkenntnis. Korrekt wäre folgende Formulierung: „Diese Struktur ist durch ihre Funktion bisher nicht (voll) verstanden und könnte eine Diskrepanz zwischen Struktur und Funktion aufweisen. Diese Diskrepanz könnte ein Hinweis auf eine abstammungsbedingte Ähnlichkeit sein.“ Die Möglichkeit der Abstammungsbedingtheit einer Ähnlichkeit ist natürlich gegeben, aber das ist nicht beweisbar. Im obigen Beispiel ist zu bedenken, dass Wale und Delphine sich teilweise anders fortbewegen als Fische; sie haben keine Bauchflossen, sondern eine Schwanzflosse (Fluke). Es kann kaum ausgeschlossen werden, dass der Skelettbau der Brustflossen der Wale und Delphine für die auszuübende Funktion optimal ist.
2. Diskrepanzen zwischen Struktur und Funktion müssen nicht notwendigerweise auf gemeinsame Abstammung hinweisen. Es gibt Fälle, in denen dies sogar unplausibel ist (s. „Design“ (https://genesis-net.de/s/0-4/)). Im Rahmen von Schöpfungsmodellen können solche Diskrepanzen durchaus plausibel gemacht werden, wenn es sie denn wirklich geben sollte „Rudimentären Organen“ (https://genesis-net.de/e/1-3-e/5-3/).
2.3 Einwand 2: Mit „Schöpfung“ kann man beliebige Ähnlichkeitsmuster oder auch die Abwesenheit von Ähnlichkeit erklären.
Dieses Argument wird von Mahner (2002) wie folgt ausgeführt: „Gewiss würde auch eine Schöpfungstheorie die abgestufte Ähnlichkeit der Arten, wie sie sich in der biologischen Systematik darstellt, erklären: Gott hat eben ähnliche Arten geschaffen. Und, so fragen die Kreationisten, ist diese Ähnlichkeit nicht gerade Beweis für die Handschrift desselben Baumeisters? Leider nein, denn eine auf Schöpfung basierende Theorie könnte auch den genau gegenteiligen Fall erklären, nämlich den, dass es keinerlei Ähnlichkeit zwischen den Arten gäbe: Gott hätte dann eben unähnliche Arten geschaffen. (In diesem Falle wäre die Idee einer Evolution wohl gar nicht erst aufgekommen.)“ Daher müsse ein „wissenschaftlicher Kreationismus“ scheitern und auf einem Schöpfungsansatz basierende Theorien könnten nicht als Alternativen zur Evolutionstheorie in Frage kommen. Kritiker wenden also ein, dass beliebige Erwartungen an das Merkmalsmuster der Lebewesen aus der Schöpfungslehre abgeleitet werden können; die Daten würden also immer „passen“.
Erwiderung: Der Einwand ist insofern berechtigt, als in der Tat keine eindeutigen Erwartungen aus der Schöpfungslehre abgeleitet werden können, doch die Situation ist bei der Evolutionslehre nicht grundlegend anders. Außerdem geht es nicht, wie Mahner schreibt, um „Beweise“, sondern um Deutungsmöglichkeiten.
Die Evolutionstheoretiker können nur bedingt und nur relativ vage bestimmte Ähnlichkeitsmuster vorhersagen; eindeutige Erwartungen können in ihrem Rahmen genauso wenig formuliert werden wie im schöpfungstheoretischen Rahmen. In der Vergangenheit wurden zwar gelegentlich bestimmte evolutionstheoretische Erwartungen an das Ähnlichkeitsmuster formuliert, doch diese bewährten sich in der Realität oft nicht, ohne dass deshalb die Evolutionslehre als Rahmenanschauung angetastet worden wäre. Die evolutions-
theoretische Praxis sieht so aus, dass zunächst nach bestimmten Kriterien Ähnlichkeitsmuster ermittelt werden, und diese im Nachhinein eine evolutionstheoretische Deutung erfahren. Im Rahmen der Schöpfungslehre wird hier methodisch nicht wesentlich anders verfahren. Auch dort können die Ähnlichkeitsmuster gedeutet und dabei auch Gesetzmäßigkeiten und weitere konkrete, widerlegbare Erwartungen formuliert werden (s. „Mosaikformen als Grundtypen und Baukastensysteme“ (https://genesis-net.de/s/0-3/3-1/)).
Wir können festhalten: Ähnlichkeiten an sich geben keine sichere Auskunft über ihre Entstehung. Dass Ähnlichkeiten für sich alleine nicht notwendigerweise eine gemeinsame Abstammung bedeuten, zeigen spezielle Ähnlichkeiten unter Menschen. Wenn sich Menschen besonders ähneln, schließen wir darauf, dass es sich um Geschwister handeln könnte. Das wäre also Abstammungsverwandtschaft: Ähnlichkeit durch Abstammung. Doch ist dies nicht sicher, denn besonders ähnliche Menschen müssen nicht unbedingt nahe verwandt sein wie sog. „Doppelgänger“ zeigen. Eine sichere Klärung bringen erst Stammbücher oder die persönliche Auskunft, also zusätzliche, andersartige Belege. Die Ähnlichkeit ist nur ein mehrdeutiges Indiz. Bei der Deutung von Ähnlichkeiten zwischen Menschenaffen und Mensch stehen diese zusätzlichen Informationen natürlich nicht zur Verfügung. In der Paläanthropologie („Urmenschenforschung“) sind solche „zusätzlichen Belegstücke“ die Fossilien, die aber wiederum kein eindeutiges Bild vermitteln. Christen sehen auch den biblischen Bericht als historisches Zeugnis an, sozusagen als geschichtswissenschaftliches Belegmaterial.
2.4 Passen verschiedene Datensätze zusammen?
Evolutionstheoretiker weisen darauf hin, dass verschiedene Datensätze (Vergleichende Biologie, Molekulare Sequenzen, Paläontologie) zusammenpassen würden, was das Gewicht der einzelnen Argumente vergrößert.
Doch die verschiedenen Datensätze passen in Wirklichkeit häufig nicht zusammen. Manchmal stimmen sie zwar ziemlich gut überein, in vielen Fällen aber nur teilweise und nicht selten sogar nur wenig. Beispielsweise legen molekulare Daten häufig frühere Divergenzzeiten nahe als die Fossilüberlieferung (Diskrepanz zwischen Molekularbiologie und Paläontologie). Die daraus resultierenden evolutionstheoretischen Probleme werden kontrovers diskutiert. Ebenso stimmen die Verwandtschaftsverhältnisse, die durch morphologisch-anatomische Daten begründet werden, oft nicht mit den molekularbiologischen Daten überein.
Im Rahmen der Evolutionslehre werden in den Fällen der Nicht-Übereinstimmung Zusatzhypothesen aufgestellt, um die ursprünglich unerwarteten Diskrepanzen zu erklären. Daher können die Nicht-Übereinstimmun-
gen der Datensätze die Evolutionslehre einerseits nicht widerlegen. Doch entspricht andererseits die Behauptung, verschiedene Datensätze würden dieselben Verwandtschaftsbeziehungen begründen, in dieser allgemeinen Form nicht den Tatsachen. Folglich bieten die verschiedenen Datensätze keine unabhängigen Belege für Makroevolution.
2.5 Konvergenzen
Die Biologen kennen zahlreiche Beispiele von tiefgreifenden Ähnlichkeiten, die nicht auf gemeinsame Abstammung zurückgeführt werden können. Die Evolutionstheoretiker unterscheiden zwischen homologen Ähnlichkeiten (Homologien) und analogen Ähnlichkeiten. Die Homologien werden auf gemeinsame Abstammung zurückgeführt, die Analogien dagegen als sog. Konvergenzen interpretiert. Konvergenz bedeutet, dass bauplanähnliche Strukturen zwei- oder mehrfach unabhängig von unterschiedlichen Ausgangsstrukturen ausgehend entstanden sind (vgl. die Beispiele in Abb. 5). Doch die Unterscheidung zwischen Homologie und Analogie (bzw. Konvergenz) ist in vielen Fällen nicht eindeutig möglich. Woran liegt das?
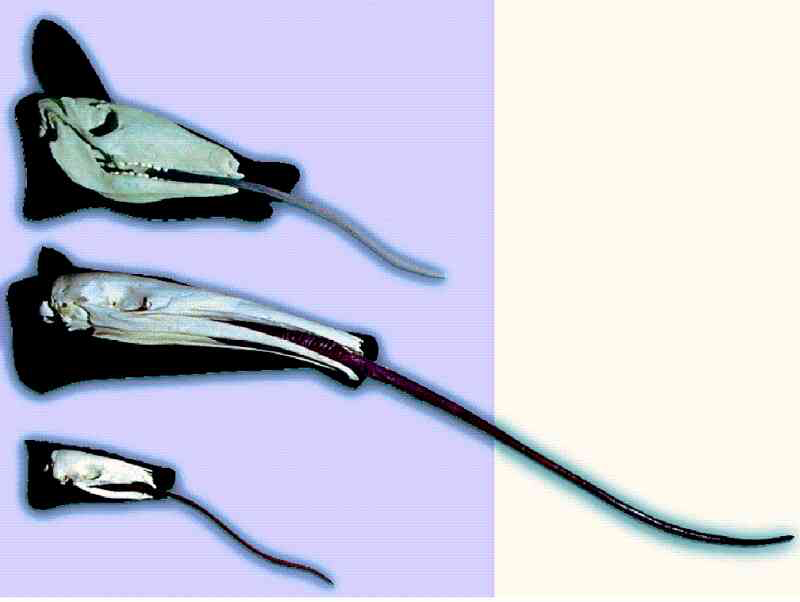
Abb. 5: Leimruten (lange, ausstülpbare, klebrige Zungen) sollen mehrfach unabhängig entstanden sein; die Ähnlichkeit gilt hier nicht als abstammungsbedingt, sondern als Konvergenz. Von oben: Erdferkel, Ameisenbär, Schuppentier. Quelle: Westfälisches Museum für Naturkunde, Münster.
Die Problematik soll an einem einfachen Beispiel klargemacht werden: Jeder wird die Pusteblume, also den Fruchtstand des Löwenzahns kennen. Die einzelnen Früchte sind als Schirmchen ausgebildet (Abb. 6 zeigt dies beim verwandten Korbblütler Wiesen-Bocksbart).

Abb. 6: Schirmchen des Wiesen-Bocksbarts (Familie Korbblütler)
Die Schirmchenkonstruktion findet sich aber nicht nur bei den Korbblütlern, zu denen der Löwenzahn gehört, sondern auch bei anderen nicht näher verwandten Arten wie z. B. den Baldriangewächsen (Abb. 7). Das heißt: Die Schirmchen müssten evolutionstheoretisch gesehen unabhängig entstanden sein, was wiederum bedeutet, dass ihre Ähnlichkeit nichts mit Abstammung zu tun hat. Abb. 8 (s. u.) verdeutlicht die Problematik für die Evolutionstheorie.

Abb. 7: Ähnlich gebaute Schirmchen des Baldrians (Familie Baldriangewächse); sie sollen unabhängig entstanden sein.
Hier wird also eine offenkundige Bauplanähnlichkeit (Schirmchenflieger) nicht als Folge einer gemeinsamen Abstammung gedeutet, sondern als mehrfacher unabhängiger evolutiver Erwerb. Würde man nämlich annehmen, dass Schirmchenflieger nur einmal entstanden sind (sich also auf einem gemeinsamen Ast des hypothetischen Stammbaums befinden), müssten dafür andere Merkmale zwei- oder mehrfach unabhängig entstanden sein.
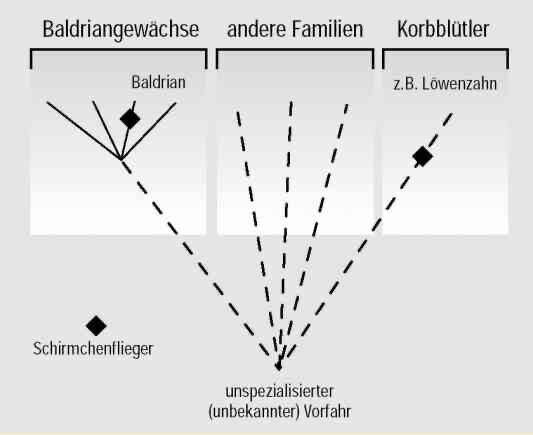
Abb. 8: Schematische Darstellung der unsystematischen Verteilung eines Bauelements im Sinne eines gedachten Baukastensystems.
Die Merkmale sind oft so verteilt, dass sie nicht in einem widerspruchsfreien Stammbaum angeordnet werden können (Abb. 8 erläutert das Grundproblem schematisch).
Merkmalsübereinstimmungen können also nicht „automatisch“ als abstammungsbedingt interpretiert werden. Wird eine Merkmalsübereinstimmung auf gemeinsame Abstammung zurückgeführt, werden sie in der Phylogenetik als Synapomorphien bezeichnet (wenn es sich zudem um sog. „abgeleitete“ Merkmale handelt). Andernfalls handelt es sich um Konvergenzen (allgemeiner: Homoplasien).
Aus dieser Situation resultieren zwei evolutionstheoretische Probleme:
1. Wie gelangt ein zielloser Prozess zwei- oder mehrfach zum selben Ergebnis? Dazu gibt es folgenden Lösungsvorschlag:
- Die übliche Antwort auf die Frage, weshalb der an sich ziellose Evolutionsprozess doch zu sehr ähnlichen Strukturen führen kann (Konvergenz), lautet: Der Evolutionsvorgang wird stark kanalisiert durch ähnliche Selektionsdrücke unter ähnlichen Umweltbedingungen. Dagegen kann aber wiederum kritisch zweierlei gesagt werden.
- Zum einen handelt es sich nur um ein theoretisches Konzept, das für größere Veränderungen (bei Makroevolution – s. den Artikel „Mikro- und Makroevolution“ (https://genesis-net.de/e/1-3-a/1-3/)) nicht empirisch belegt ist. Empirisch nachvollziehbar sind Konvergenzen im mikroevolutiven Rahmen. Gleiche Selektionsdrücke können auf der Basis einer polyvalenten Stammform zu ähnlichen Spezialisierungen führen, doch führen solche Vorgänge nicht zur Entstehung neuer Baupläne oder Bauteile, sondern zu immer stärkerer Spezialisierung (Selektion).
- Zum anderen kommen Konvergenzen in vielen Fällen vor, in denen überhaupt kein Zusammenhang mit gleichartigen Selektionswirkungen bzw. gleichartigen Umweltbedingungen erkennbar ist. Details zu diesem Argument finden Sie im PDF zu diesem Artikel, dort im Abschnitt „Lösungsversuche“: https://www.wort-und-wissen.org/wp-content/uploads/rudiment_lk2.pdf.
2. Wie können Homologien und Konvergenzen unterschieden werden? (Vorgehensweise in der Phylogenetik).
- Wie kann man überhaupt Stammbäume rekonstruieren (Phylogenetik)? Es ist ja oft nicht eindeutig bestimmbar, welche Ähnlichkeiten homolog (im Sinne von „abstammungsbedingt“ = Synapomorphien) sind und welche nicht. Aber nur die abstammungsbedingten Ähnlichkeiten dürfen in der Phylogenetik zugrunde gelegt werden. Nähere Ausführungen dazu finden sich in der zu diesem Artikel: https://www.wort-und-wissen.org/wp-content/uploads/rudiment_lk2.pdf.
Das Phänomen der Konvergenz ist keine Randerscheinung, mit der es die Biologen nur in seltenen Sonderfällen zu tun hätten, sondern es ist weit verbreitet. Im Folgenden Abschnitt kommen dazu einige Beispiele.
2.6 Baukastensystem
Das Konvergenzproblem stellt sich noch in vielen anderen Fällen, auch bei wesentlich komplexeren Strukturen. Wir nennen nachfolgend einige Beispiele, um einen Eindruck zu vermitteln. Es geht dabei nicht um offen-
kundige Fälle von Analogien wie Insekten- und Vogelflügel; denn hierbei handelt es sich unumstritten um grundverschiedene Baupläne. Es geht vielmehr um solche Beispiele, bei denen man nach den gebräuchlichen Homologiekriterien zunächst annehmen würde, es handle sich um Homologien (wie bei den Schirmchenfliegern im oben genannten Beispiel).
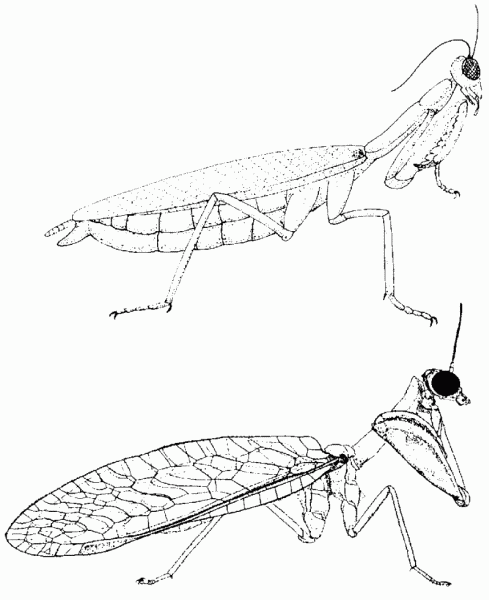
Abb. 9: Frappierend ähnlich gebaut und doch nicht abstammungsverwandt: Fangschrecke (Mantis religiosa, oben) und Fanghafte Mantispa styriaca. Quelle: Nach Sudhaus & Rehfeld 1992.
Einige besonders augenfällige Beispiele sind die Warmblütigkeit bei Vögeln und Säugetieren (sie soll unabhängig erworben worden sein), Fangbeine bei Fangschrecken und Netzflüglern (Abb. 9), das Vorkommen eines Brustbeinkiels bei Vögeln, Flugsauriern und Fledermäusen (Abb. 10), pneumatisierte (lufthaltige) Knochen bei Flugsauriern und Vögeln, malpighische Gefäße (Ausscheidungsorgane) bei Arachniden und Antennaten.
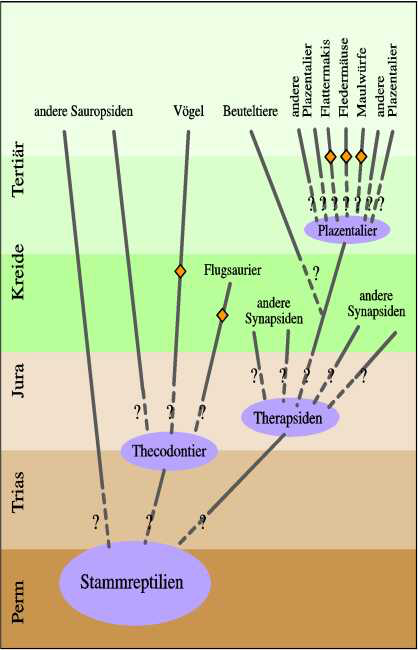
Abb. 10: Vereinfachter Stammbaum mit der Position der Organismengruppen, die einen Brustbeinkiel besitzen (mit Raute markiert).
Fleischfressende Pflanzen verteilen sich auf ganz unterschiedliche Familien, ebenso CAM-Pflanzen (Pflanzen mit speziellen Stoffwechselfähigkeiten zur effektiven Nutzung von CO2) und sukkulente Pflanzen. Weiter können zahlreiche Bestäubungs- und Verbreitungsmechanismen von Samen und Früchten genannt werden usw. Der bekannte Botaniker Cronquist (1987, 24) stellt fest, dass die Angiospermen (bedecktsamige Blütenpflanzen) mit Konvergenzen durchsetzt sind.
Moore & Willmer (1997) zeigen in einer Überblicksarbeit auf, dass auch bei den Wirbellosen Konvergenzen weit verbreitet sind und dass das Ausmaß an Konvergenzen stark unterschätzt worden ist. Beispielsweise muss angenommen werden, dass die Tracheen (von außen in den Körper sich verzweigende Luftröhren zur Atmung) mindestens viermal unabhängig entstanden sind, wobei in den betroffenen Tieren das Tracheensystem in fast identischer Weise gebaut ist (Moore & Willmer 1997, 14). Bei den Wirbellosen beschränken sich Konvergenzen zudem nicht vornehmlich auf „untergeordnete Merkmale“; sogar komplexe „Design“-Merkmale sind oft konvergent und für die Rekonstruktion der Phylogenese irreführend (Moore & Willmer 1997, 16).
Ein besonders interessantes Beispiel sind Leimruten: Das sind lange, ausstülpbare, klebrige Zungen. Ein solches Organ soll im Laufe der angenommenen Evolution fünfmal unabhängig entstanden sein, nämlich beim Ameisenbär, Schuppentier, Erdferkel, Specht und Chamäleon (Abb. 5). Mit den „Leimruten“-Zungen werden kleine Insekten (vornehmlich Ameisen oder Termiten) aufgenommen.
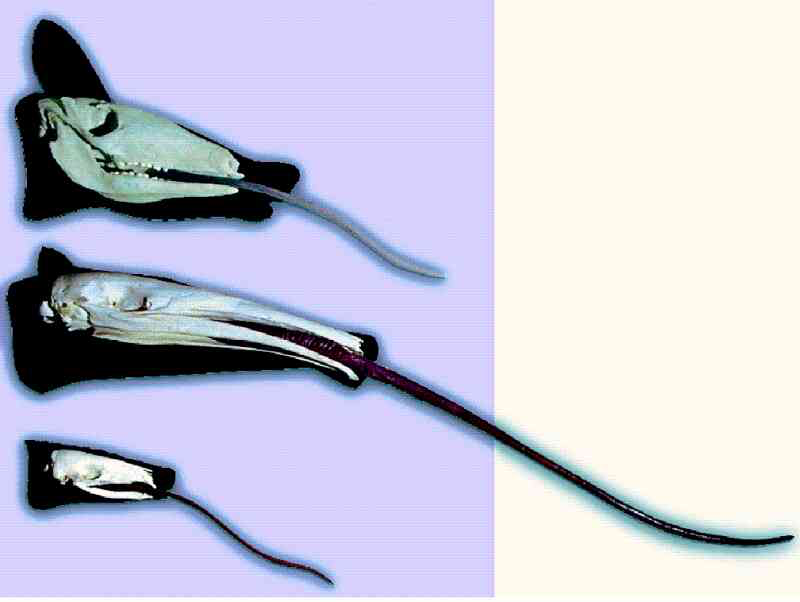
Abb. 5: Leimruten (lange, ausstülpbare, klebrige Zungen) sollen mehrfach unabhängig entstanden sein; die Ähnlichkeit gilt hier nicht als abstammungsbedingt, sondern als Konvergenz. Von oben: Erdferkel, Ameisenbär, Schuppentier. Quelle: Westfälisches Museum für Naturkunde, Münster.
Im Einzelnen gibt es zwar mehr oder weniger große Unterschiede im Bau der einzelnen Leimruten, doch insbesondere bei den drei Säugergruppen (Ameisenbär, Schuppentier und Erdferkel) und beim Specht einige tiefgreifende Gemeinsamkeiten: Der Kiefer ist schmal und zahnlos (oder die Zähne sind nur schwach ausgebildet), die Schnauze ist verlängert und die Mundöffnung verengt. Die Zunge ist sehr lang (bis zu 60 cm) und klebrig. Für ihre Betätigung werden besondere Muskeln benötigt. Als Kauapparat dient der Magen, der als Kaumagen ausgebildet ist. Das heißt: die aufgenommene Nahrung wird im Magen z. B. durch Platten und Muskeltätigkeit mechanisch zerrieben und ausgepresst. Insgesamt handelt es sich um eine komplizierte Apparatur. Wie sie einmalig durch evolutionäre Zufalls- und Auslesevorgänge (Mutation und Selektion) entstanden sein sollte, ist schon unklar. Eine mehrmalige Entstehung einer solch komplizierten Konstruktion auf Zufallswegen ist noch viel unglaubhafter. Dieses Beispiel zeigt darüber hinaus, dass aufgrund von Ähnlichkeit gar nicht automatisch auf gemeinsame Abstammung geschlossen werden kann. Denn die Tiere, die Leimruten besitzen, sind sich in anderen Merkmalen insgesamt so unähnlich, dass sie auf ganz verschiedene Äste des evolutionären Stammbaums gesetzt werden.
Es zeigt sich also folgende Situation: Die Merkmale sind in vielen Fällen so unsystematisch verteilt, dass sie in den einzelnen Arten bzw. höheren Taxa baukastenartig zusammengesetzt erscheinen. Es fällt auf, dass Taxonomen immer wieder den Begriff einer „Toolbox“ zur Beschreibung der Merkmalsverteilung verwenden. Im Rahmen eines Baukastensystems sind zunächst beliebige Merkmalskombinationen denkbar. (Allerdings kann es durchaus auch Einschränkungen der Kombinationsmöglichkeiten geben, wenn bestimmte Merkmalskombinationen funktionell oder ökologisch nicht zusammenpassen.)
Evolutionstheoretisch lässt sich ein solches Baukastensystem nicht leicht erklären, weil eine größere Zahl von Konvergenzen angenommen werden muss. Die problematische Annahme von Konvergenzen könnte vermieden werden, wenn das Auftreten baugleicher Merkmale durch Gentransfer plausibel gemacht werden könnte, doch ist diese Möglichkeit derzeit spekulativ (s. Artikel „Gene tinkering“ (https://genesis-net.de/e/1-3-c/3-4/). Im Rahmen eines Schöpfungsmodells ist eine baukastenartige Verteilung von Merkmalen verständlich (genauere Begründung im Artikel „Mosaikformen“ (https://genesis-net.de/s/0-3/3-1/); zur Vertiefung: PDF zu diesem Artikel: https://www.wort-und-wissen.org/wp-content/uploads/rudiment_lk2.pdf)
2.7 Schlussfolgerungen
Ähnlichkeiten können nach bestimmten biologischen Kriterien (Homologiekriterien) als „homolog“ eingestuft werden. Das ist unabhängig von der Vorgabe der Evolutionstheorie oder einer anderen Ursprungstheorie möglich. Evolutionstheoretisch werden manche, und eben nur manche Homologien (Synapomorphien) als abstammungsbedingte Ähnlichkeiten interpretiert. Es zeigt sich nämlich, dass unterschiedliche homologe Merkmale in vielen Fällen verschiedene, sich widersprechende Abstammungsverhältnisse ergeben. Daher können nicht alle nach den Homologiekriterien ermittelten Homologien auf gemeinsame Abstammung zurückgeführt werden. Vielmehr muss ein Teil der so bestimmten Homologien zwei- oder mehrfach unabhängig entstanden sein: Konvergenzen. Doch welche sind das? Durch Algorithmen wird festgestellt, welche Stammbaumvariante mit der geringsten Anzahl an Konvergenzen auskommt. Aus diesem Stammbaum ergibt sich dann, welche Merkmalsübereinstimmung als abstammungsbedingt gelten können (= Synapomorphien in der Terminologie des Cladismus).
Folglich kann erst bei Vorgabe eines Stammbaumschemas bestimmt werden, welche Merkmalsüberseinstimmung synapomorph, also abstammungsbedingt sind. Wenn aber ein Stammbaumschema vorgegeben werden muss, um abstammungsbedingte Homologien bestimmen zu können, ist es umgekehrt nicht möglich, aus Homologien auf Abstammung zu schließen.
Das heißt: Die Beobachtungsdaten (der Befund der Ähnlichkeit) werden nachträglich in das vorgegebene Evolutionskonzept eingebaut. Die Daten selber „produzieren“ oder erzwingen das Deutungsschema nicht, sondern dieses wird vorgegeben. Gleichgültig, welches Ursprungskonzept zugrunde gelegt wird, kann nur das Ergebnis der Entstehung untersucht werden, nicht aber der Weg zu ihrer Entstehung (vgl. Abb. 3). Dieser Weg muss hypothetisch vorgegeben werden. Seine Plausibilität kann dann anhand der Beobachtungsdaten eingeschätzt werden.
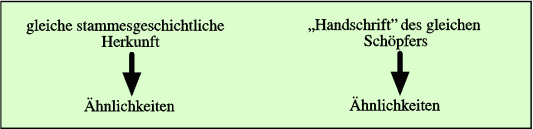
Abb. 3: Zwei mögliche Begründungen für das Vorliegen von Ähnlichkeiten.
Noch einmal anders formuliert: Es muss ein Abstammungsschema vorgegeben werden, aufgrund dessen dann manche Ähnlichkeiten als homolog im Sinne von abstammungsbedingt (synapomorph) interpretiert werden können. Wenn aber Evolution vorausgesetzt werden muss, liegt an dieser Stelle kein unabhängiger Beleg für Evolution vor.
Weitere Details finden sich hier im wissenschaftlichen Aufsatz zum Thema: https://www.wort-und-wissen.org/wp-content/uploads/rudiment_lk2.pdf
2.8 Zitierte Literatur
Cronquist A (1987) A botanical critique of cladism. Bot. Rev. 53, 1-52.
Mahner M (2002) Kreationismus. In: Sauermost R und Freudig D (Red.) Lexikon der Biologie, Bd. 8. Heidelberg, S. 202-203.
Sudhaus W & Rehfeld K (1992) Einführung in die Phylogenetik und Systematik. Stuttgart, Jena, New York.
Moore J & Willmer P (1997) Convergent evolution in invertebrates. Biol. Rev. 72, 1-60.
2.9 Weiterführende Literatur
R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001, Kap. V.9 und VII.17.4.
R. Junker: Ähnlichkeiten, Rudimente, Atavismen. Design-Fehler oder Design-Signale? Studium Integrale. Holzgerlingen, 2002. (U. a. bietet dieses Buch eine detaillierte Analyse des Ähnlichkeits-Arguments der Evolutionstheorie).
Autor: Reinhard Junker, 10.01.2004
Aktualisiert am 07.01.2024 (B. Scholl); © beim Autor; alter Link: 2004, https://www.genesisnet.info/schoepfung_evolution/e41301.php