Artbildung
Durch räumliche Trennung können Teilpopulationen einer Art unter verschiedene Auslesebedingungen geraten, es können verschiedene Mutationen und Rekombinationen eintreten. Dies führt zu einem Verschiedenwerden, das bis zur Entstehung von Kreuzungsbarrieren führen kann, was gleichbedeutend mit einer Artbildung ist. Solche Vorgänge führen zu Spezialisierungen, nicht zu Höherentwicklung und sind mikroevolutiver Natur.
1.0 Inhalt
„Mutation“ (https://genesis-net.de/e/1-3-b/2-1/) und „Selektion“ (https://genesis-net.de/e/1-3-b/2-2/) sind grundlegende Mikroevolutionsvorgänge, die zu einem Wandel der Arten führen. Dadurch vergrößert sich jedoch die Anzahl der Arten nicht, die Arten ändern sich nur. Evolution beinhaltet aber nicht nur Wandel im Laufe der Zeit, sondern auch Entstehung von Vielfalt, eine Zunahme der Anzahl der Arten. Entsprechend ist neben dem Artenwandel die Artbildung ein zweiter elementarer evolutionärer Vorgang.
In diesem Artikel wird erklärt, unter welchen Bedingungen Artbildung erfolgen kann, welche Vorgänge dabei ablaufen und in welcher Geschwindigkeit dies möglich ist.
1.1 Was ist Artbildung?
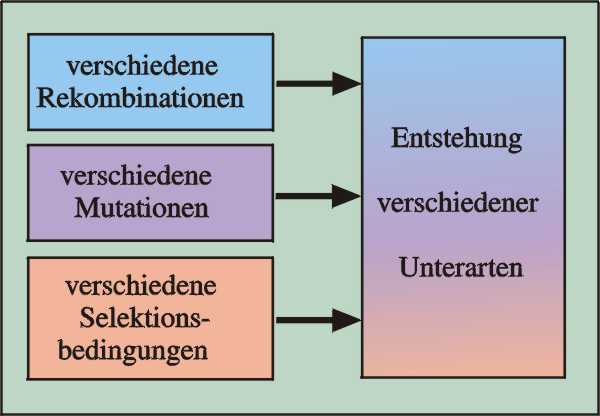
Abb. 66: Die unterschiedliche Wirkung der Evolutionsfaktoren in getrennten Teilpopulationen.
Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
Unter Artbildung versteht man die Aufspaltung von einer Art in zwei oder mehrere Arten. Voraussetzung für Artbildung ist eine geographische oder ökologische Trennung der Ausgangsart. In den geteilten Populationen können die Evolutionsfaktoren Mutation, Selektion und Rekombination unterschiedlich wirken (Abb. 66). Wenn bei späterer Wiederüberlappung der Besiedlungsgebiete keine Kreuzungen mehr eintreten, ist die vormalige Art in zwei Arten aufgeteilt worden (vgl. Abb. 67).
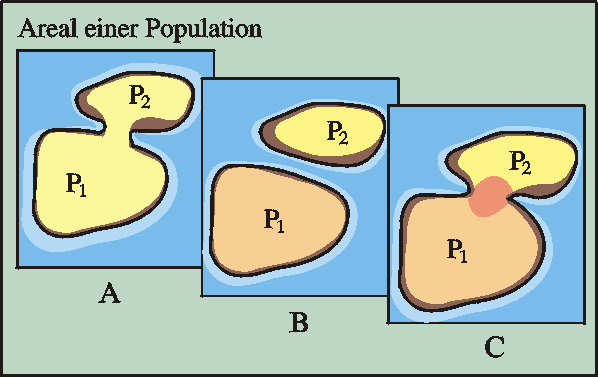
Abb. 67: Räumliche Trennung (Separation) einer Population, anschließende Divergenz (Mitte) und schließlich genetische Isolation, d. h. Aufspaltung in zwei Arten. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
Wir betrachten zunächst ein gut belegtes Beispiel. Neue Arten entstanden in Südengland auf Böden, die durch Schwermetalle aus Bergwerkshalden vergiftet waren. Abb. 68 zeigt, was sich dort ereignet hat. Die schwermetallverseuchten Bergwerkshalden sind derart belastet, dass angewehte Samen normalerweise nicht auskeimen können oder sehr schnell absterben (†). Einige wenige wachsen jedoch aus und können fruchten. Diese gründen eine neue Population an einem neuen Standort. Ihre Nachkommen sind alle in der Lage, die vergifteten Halden zu besiedeln. Es hat sich herausgestellt, dass die Haldenpflanzen sich kaum noch oder gar nicht mehr mit den nächstverwandten Formen umliegender unbelasteter Standorte, von denen sie stammen, kreuzen. Damit ist eine genetische Isolation eingetreten. Das heißt nach dem Biospezies-Kriterium, dass neue biologische Arten entstanden sind (s. Artikel „Artbegriffe“ (https://genesis-net.de/e/1-3-a/1-4/)). Biologische Arten entstehen also auch heute unter natürlichen Bedingungen. Liegt damit aber auch ein Hinweis auf beginnende Höherentwicklung (Makroevolution) vor? Dieser Frage stellt sich der folgende Abschnitt.
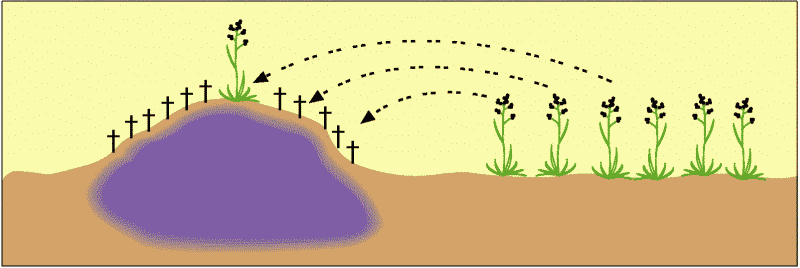
Abb. 68: Bergwerkshaldenpflanzen. Die meisten Samen, die auf die giftigen Böden der Bergwerkshalden fallen, keimen nicht, oder die Pflanzen sterben nach Keimung ab (†). Nur wenige Vorangepasste können auswachsen. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
1.2 Was geschieht bei der Artbildung?
Die oben erwähnten Bergwerkshaldenpflanzen wurden genetisch untersucht. Es stellte sich heraus, dass die Pflanzen, die auf den giftbelasteten Böden wachsen, keine neuen Eigenschaften erworben haben, sondern dass die Fähigkeit der Gifttoleranz bereits in den jeweiligen Arten vorhanden war, bevor sie die Giftböden besiedelten. Nur einige solcher Arten, von denen ein Teil auch auf den normalen Standorten gifttolerant ist, konnten die Giftböden besiedeln. Die Gifttoleranz gehört bei diesen Arten zur natürlichen Variabilität. (Der Anteil der gifttoleranten Arten ist so hoch, dass er bei weitem nicht durch spontane Mutationen erklärbar ist.)
Offenbar erfolgte eine Auslese der zuvor schon gifttoleranten Formen. Die einseitige Auslese hat eine Spezialisierung zur Folge (vgl. Abb. 69). Damit gekoppelt ist z. T. eine Verringerung der Kreuzbarkeit mit den Normalformen der unbelasteten Böden, womit genetische Isolation eintritt und eine Artbildung erfolgt. Die genetische Isolation kann in diesem Fall z. B. durch eine Verschiebung der Blühzeiten eintreten.
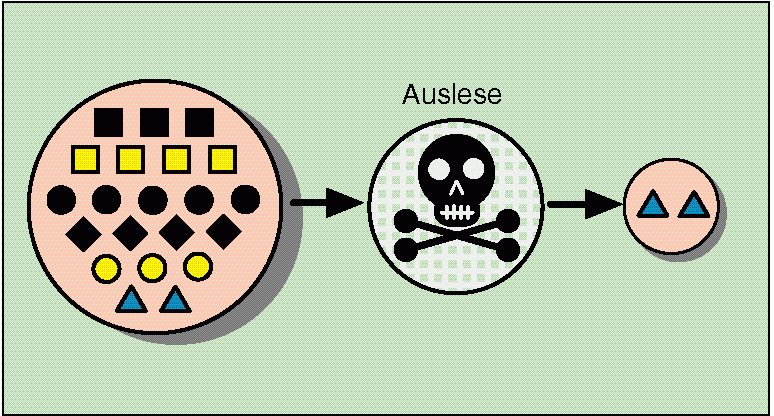
Abb. 69: Extreme Selektion auf Giftböden: Unter einer großen Anzahl verschiedener genetischer Varianten, die verschieden ausgeprägte Fähigkeit der Gifttoleranz haben, überlebt nur eine Variante. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
Die Gifttoleranz rührt übrigens wahrscheinlich daher, dass die Aufnahme von Mineralsalzen aus dem Boden eingeschränkt ist; es handelt sich also nicht um einen evolutionären Fortschritt, sondern um eine Einschränkung, die sich in der speziellen Umgebung, auf den Giftböden, als vorteilhaft erweist.
Die Vorgänge bei den Bergwerkshaldenpflanzen zeigen wie ähnlich gelagerte Fälle, dass Artbildung nicht als beginnende Höherentwicklung anzusehen ist, sondern oft mit Spezialisierung und damit mit Verarmung des Genpools verbunden ist. Mehrfach aufeinanderfolgende Artaufspaltungen führen daher tendenziell zu einer Verarmung des Genpools und damit zu einer Verminderung der Variabilität (vgl. Abb. 70). Zwar können Mutationen der Verarmung des Genpools entgegenwirken, doch zeigen Studien, dass der Trend zur Verarmung damit nicht gebrochen wird. Wenn die aufgespaltenen Arten dann aufgrund ihrer einseitigen Spezialisierung nicht mehr flexibel auf Umweltveränderungen reagieren können, sind sie vom Aussterben bedroht.
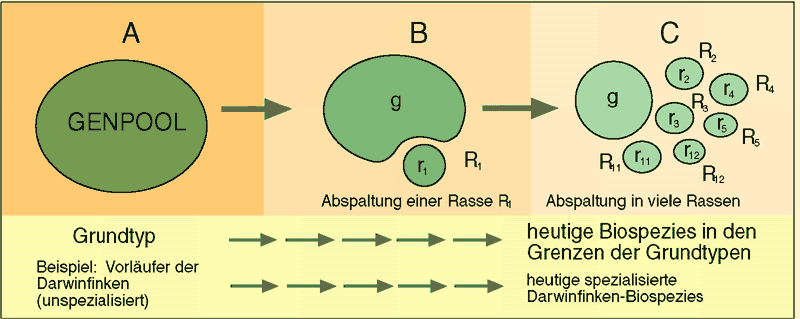
Abb. 70: Hintereinander erfolgende Abspaltungen von Teilpopulationen (Rn) mit dem Genpool rn führen zu Spezialisierungen und Genpoolverarmung, was bildlich durch die kleiner werdenden Ovale angedeutet wird. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001
Wir können festhalten: Die Entstehung neuer biologischer Arten ist ein mikro-
evolutiver Vorgang und gibt keine Hinweise auf Makroevolution.
1.3 Gründereffekt, Gendrift und sympatrische Artbildung
Nach dem klassischen Artbildungskonzept ist für die Abspaltung einer neuen Art eine geographische Trennung erforderlich. Eine Gründergruppe muss von der Population abgetrennt werden: Gründereffekt. Je kleiner die Gründergruppe ist, desto eher weicht ihr Genpool [= alle Allele der Gene einer Art] vom Durchschnitt des Gesamtgenpools ab. Viele Gene sind im Genpool in mehreren Allelen [= Zustandsformen eines Gens] vertreten. In einer kleinen Gründergruppe bleibt davon unter Umständen nur ein kleiner, nichtrepräsentativer Teil übrig. Eine solche Verschiebung der Allelhäufigkeiten, die nicht durch Selektion hervorgerufen wurde, sondern zufällig ist, nennt man Gendrift. Die Gründergruppe startet also oft von einer anderen genetischen Ausgangsbasis als der Rest der Population. Damit ist vom Start weg bereits eine Voraussetzung für die Artspaltung gegeben. Weitere Mechanismen wie beschrieben können zu weiteren mikroevolutiven Änderungen bis hin zu Kreuzungsbarrieren führen.
Es gibt zunehmend Befunde, die eine Artbildung auch ohne räumliche Trennung wahrscheinlich machen. Man nennt diesen Vorgang sympatrische Artbildung (sympatrisch = „im selben Gebiet“). Wenn beispielsweise Parasiten, die auf einen bestimmten Wirt spezialisiert sind, plötzlich ihren Wirt dauerhaft wechseln, sind sie und ihre Nachkommen von der Ausgangsgruppe ökologisch getrennt und können sich isoliert voneinander (mikroevolutiv) weiterentwickeln.
1.4 Geschwindigkeit der Artbildung
Eine Reihe von Studien hat gezeigt, dass Artbildung innerhalb weniger Generationen erfolgen kann, wenn die Ausgangsgruppe genetisch polyvalent (vielseitig, variabel) und die Gründergruppe sehr klein und damit genetisch „einseitig“ ist. Der Genpool einer kleinen Gründerpopulation oder einer kleinen Restpopulation kann am leichtesten eine gründliche und dauerhafte Änderung erfahren. Diesem Zusammenhang liegen zahlreiche Beobachtungen zugrunde. Es sei an das Beispiel der Bergwerkshaldenpflanzen erinnert. Die auf den neuartigen Standorten der giftbelasteten Bergwerkshalden mehrfach beobachtete Neuentstehung von Biospezies vollzog sich offenbar während weniger Generationen. Hier ist aufgrund einer gravierend veränderten Umwelt ein Großteil der Population der Selektion zum Opfer gefallen.
Das nachfolgende Beispiel zeigt, dass auch morphologische Änderungen in kürzester Zeit zu neuen Biospezies führen können: Bei der Clarkie wurde eine Verlust-Mutante bicalyx beobachtet, bei der die Blütenblätter in kelchblattartige Gebilde umgewandelt wurden (Abb. 71; es handelt sich um eine sog. homeotische Mutation). Diese Mutante hat sich im Freiland stabil etabliert. Bei diesem Grundtyp wurden wiederholt auch andere Mutationen beobachtet, welche die Fruchtbarkeit zwischen Mutanten und Eltern deutlich herabsetzen – ein Schritt hin zur Artaufspaltung. In Kombination mit der bicalyx-Mutante könnte dies leicht zu einer neuen Biospezies führen, die sich äußerlich deutlich von der Elternart unterscheiden würde.

Abb. 71: Bicalyx-Mutante der Clarkie mit doppelter Ausbildung des Kelchblattkreises auf Kosten des Kronblattkreises. Quelle: Aktualisiert am 07.01.2024 (B. Scholl); © beim Autor; alter Link: Leslie D. Gottlieb.
Artbildung durch Einschränkung des Modifikationspotentials. Dieses wenig beachtete Modell zur Artbildung liefert weitere Hinweise auf die Möglichkeit schneller Artbildung. Es geht von der Annahme aus, dass die Stammformen der heutigen Biospezies ein relativ großes Modifikationsspektrum (durch Umweltreize auslösbare Anpassungsmöglichkeiten) besaßen, welches im Laufe der Zeit mehr oder weniger stark durch Defektmutationen oder Gründerereignisse eingeschränkt wurde (Abb. 72). Durch Einengung der ursprünglichen Modifikationsspannen entstehen spezialisierte biologische Arten. Dies äußert sich z. B. darin, dass die Nachfahren im Vergleich zu den Vorläufern, die ein weites Spektrum möglicher Standorte hatten, nur relativ spezielle Standorte besiedeln können. Dieser von Lönnig (1993) entwickelte Ansatz ist vor allem im Rahmen des Grundtypkonzepts der Schöpfungslehre interessant und wird im Fachgebiet „Grundtypenbiologie“ (https://genesis-net.de/s/0-3/) näher erläutert. Da die Artbildung in diesem Fall auf Verlusten beruht, kann sie u. U. in kurzen Zeiträumen erfolgen.
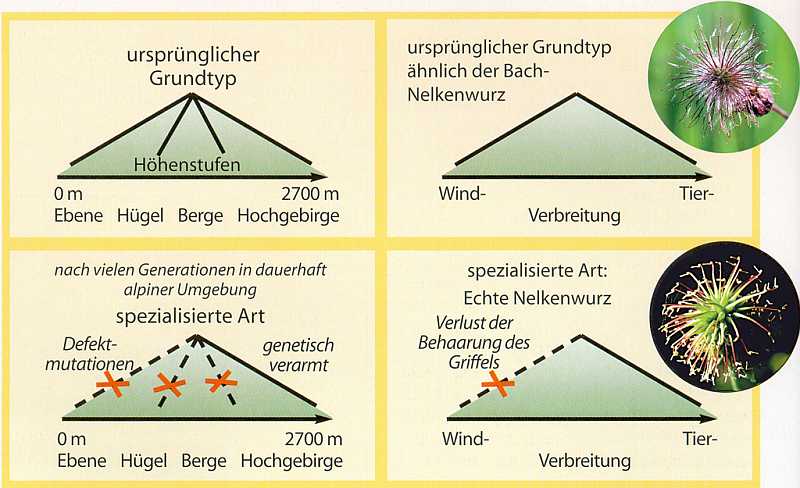
Abb. 72: Links: Modifikationspotential von Grundtypen (oben), beispielhaft bezogen auf die Höhenverteilung und Einschränkung des Potentials bei Pflanzen, die nur auf der alpinen Stufe leben können (unten). Rechts: Ein Beispiel liefern Nelkenwurzarten. Die Bach-Nelkenwurz (oben) besitzt Früchte, die (noch?) relativ vielseitig verbreitet werden können: sowohl durch den Wind (aufgrund der Behaarung), durch Tiere (mit Hilfe des Hakens) und durch Verkleben mittels Wasserhaftung. Damit korrespondiert eine weite Verbreitung von Tallagen bis weit über 2000 m Höhe auf Bergwiesen. Die Echte Nelkenwurz (unten) besitzt dagegen einen nur schwach behaarten Hakengriffel, der nur Tierverbreitung (Klettverbreitung) erlaubt. Entsprechend kommt diese Art nur in tieferen Lagen, an Waldrändern, in Gärten usw. vor. Diese Art ist stärker spezialisiert. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
1.5 Zusammenfassung
Artaufspaltungen sind nicht als erste Schritte einer Höherentwicklung zu werten. Nachweisbar sind nur mikroevolutive Vorgänge. Empirische Befunde deuten darauf hin, dass Artaufspaltungen zu Genpoolverarmung, zu Spezialisierung und in Entwicklungssackgassen (erhöhte Aussterbegefahr) führen und nicht den Beginn weiterreichender Abwandlungen darstellen.
Autor: Reinhard Junker, 01.01.2004
Aktualisiert am 07.01.2024 (B. Scholl); © beim Autor; alter Link: 2004, https://www.genesisnet.info/schoepfung_evolution/i41244.php