Modell für einen Umbruch in der Schöpfung
Der Übergang von einer Ursprungsökologie ohne Tod in der Schöpfung in die heutige Ökologie kann in vielen Fällen nur umbruchsartig erfolgt sein. Dieser Vorgang ist unzugänglich und daher unerforschbar. Die heutige Biologie liefert aber Modelle dafür, wie man sich einen solchen Umbruch vorstellen kann, ohne dass eine Neuschöpfung erforderlich ist und ohne dass destruktive Strukturen in der ursprünglichen Schöpfung bereits verwirklicht gewesen sein mussten.
1.0 Inhalt
In diesem Artikel wird ein Modell vorgestellt, wie ein abrupter Übergang von einer ursprünglichen Schöpfung ohne destruktive („fallsgestaltige“) Räuber-Beute-Beziehungen und ohne Tod in die heutige Ökologie des Fressens und Gefressenwerdens denkbar sein könnte.
1.1 Problemstellung
Im Artikel „Biblische Aussagen zur Existenzweise der Lebewesen“ (https://genesis-net.de/s/0-5/2-2/) wurde die biblische Sicht von den destruktiven Seiten der Schöpfung dargelegt. Die Bibel charakterisiert diese Seite der Schöpfung als Ausdruck eines kräftigen Missklangs, nicht als Selbstverständlichkeit. Dieser Missklang gehört aber nicht zur ursprünglichen, „sehr guten“ Schöpfung, sondern ist erst nachträglich in die Schöpfung eingedrungen. Da der Sündenfall des Menschen das Einfallstor für den Tod in der Schöpfung war, kann man die Strukturen in der Schöpfung, die mit dem Fressen und Gefressenwerden von Tieren zu tun haben, auch als „fallsgestaltig“ bezeichnen, um den Ausdruck „destruktiv“ zu vermeiden, der zu kurz greift. Mit „fallsgestaltig“ sind also Lebensstrukturen gemeint, die zum Erbeuten und Verzehr von Tieren benötigt werden, ferner Einrichtungen für Parasitismus, aber auch Mechanismen der Feindabwehr. Fallsgestaltige Strukturen werden detailliert im Artikel „Todesstrukturen in der Schöpfung“ (https://genesis-net.de/s/0-5/2-1/) beschrieben.
Vor dem biblischen Hintergrund stellt sich die Frage, wie man sich einen Umbruch von der ursprünglichen in die jetztzeitliche Schöpfung biologisch denken kann, wenn man nicht annehmen will, dass in der „sehr guten“ Schöpfung bereits die fallsgestaltigen Kennzeichen angelegt waren. Wie kamen die fallsgestaltigen Strukturen in die Schöpfung? Da 1. Mose 1-11 ein historisches Geschehen schildert (vgl. „Die Bindung der Erdgeschichte an den Sündenfall des Menschen“, https://genesis-net.de/s/0-2/1-3/), hat es einen Umbruch von einer ursprünglich „sehr guten“ (1. Mose 1,31) zu einer „völlig verderbten“ (1. Mose 6,12) Schöpfung gegeben. Was hat zu diesem Umbruch geführt? Welche Veränderungen haben ihn herbeigeführt? Was ist im Einzelnen dabei geschehen? Sind solche Fragen überhaupt beantwortbar?
Diese Fragen stellen sich im Rahmen der Evolutionslehre nicht. Während einer allgemeinen Evolution der Lebewesen kann es keinen Umbruch zwischen den Lebensbedingungen vor und nach einem Sündenfall gegeben haben. Doch die evolutionäre Weltsicht steht im Widerspruch zur biblischen Sicht von der Schöpfung (vgl. „Die biblische Urgeschichte im Neuen Testament“ (https://genesis-net.de/s/0-5/1-2/)). Daher wird hier eine andere Lösung verfolgt.
1.2 Was muss sich bei einem Umbruch in der Schöpfung geändert haben?
Wird 1. Mose 1-11 also historisch-faktisch verstanden, erhebt sich die Frage, wie ein Umbruch von der Ursprungsökologie (ohne Tod) in die heutige Ökologie (mit Tod) vonstatten gegangen sein könnte. Es soll ausgeschlossen werden, dass Gott Fallsgestaltiges am Anfang geschaffen und damit von vornherein gewollt hat, denn dies bedeutete, dass es zur Schöpfung wesensmäßig gehörte. Das heißt: Alle Strukturen und Verhaltensweisen, die ausschließlich zum Finden, Erbeuten, Verzehren und Verdauen lebendiger tierischer Nahrung benötigt werden, haben in der ursprünglichen Schöpfung keinen Platz (fallsgestaltige Strukturen). Im Artikel „Todesstrukturen in der Schöpfung“ (https://genesis-net.de/s/0-5/2-1/) wird einiges dazu beschrieben.
Da die Unterschiede zwischen einer Ursprungs- und der Rezent-Ökologie tiefgreifend sein müssen, erscheint es unmöglich, den Aufbau und die Gesetze der Biosphäre vor dem Sündenfall anschaulich zu beschreiben. Wir können uns keine Begriffe von einem Ökosystem machen, in dem die in 1. Mose 1,30 angedeuteten Zustände herrschen (keine Tiernahrung). Die ursprüngliche Schöpfung vor dem Fall ist und bleibt ein echtes Geheimnis. Daher ist es auch nicht möglich, einen Übergang von der ursprünglichen zur heutigen Ökologie auszumalen. Der nachfolgende spekulative Deutungsversuch soll daher nur als Denkhilfe verstanden werden, nicht jedoch als Versuch, etwas gedanklich in den Griff zu bekommen, was jenseits des Begreifens und der Begriffsmöglichkeiten des Menschen liegt.
1.3 Entwicklung zur fallsgestaltigen Lebensweise?
Ein aus biologischen Gründen nur in manchen Fällen vertretbarer Deutungsversuch ist die Annahme einer allmählichen Entstehung der Raubtiergestalt bzw. der räuberischen Lebensweise durch mikroevolutive, empirisch bekannte Prozesse nach dem Sündenfall. (Zum Begriff „Mikroevolution“ siehe „Mikro- und Makroevolution“ (https://genesis-net.de/e/1-3-a/1-3/).) In vielen Fällen reicht dies aber nicht aus, nämlich dann, wenn die Entstehung fallsgestaltiger Strukturen Makroevolution erfordert.
Die Vorstellungen eines allmählichen Umbruchs scheitert außerdem an der Tatsache, dass alle Lebewesen ökologisch durch Fressen und Gefressenwerden miteinander verbunden sind. Ein allmählicher Übergang von einem ganz anders organisierten Ökosystem in heutige komplizierte Bedingungen entzieht sich ebenfalls einer Erklärung durch bekannte biologische Prozesse.
1.4 Sprunghafte Veränderungen?
Der nachfolgende Deutungsversuch nimmt auf die Erkenntnis der Biologie Bezug, dass die Lebensprozesse hierarchisch organisierten Steuerungen unterliegen. Schon im Erbgut muss zwischen Strukturgenen und Regulatorgenen unterschieden werden. Die Strukturgene stehen für Eiweiße (Bau- oder Stoffwechselproteine), die Regulatorgene für solche Proteine, die beim kontrollierten Ablesen der Strukturgene benötigt werden. Doch die Regulatorgene steuern selber nicht. Sie sind lediglich eine notwendige Voraussetzung dafür, dass die Information auf den Strukturgenen kontrolliert abgelesen und für den Zellstoffwechsel nutzbar gemacht werden kann. Die Regulatorgene bedürfen ihrerseits der Regulation. Dies geschieht z. B. durch Hormone, das sind Botenstoffe, die über die Blutbahn zirkulieren. Diese aber wiederum müssen selber von einer noch höheren Instanz reguliert werden und so weiter. Man kann also festzuhalten, dass man nach bisherigen Forschungsergebnissen davon ausgehen muss, dass das Erbgut und die vorgeschalteten Steuerungen hierarchisch strukturiert sind. (Das ist stark vereinfacht dargestellt; die tatsächliche Situation ist durch Rückkopplungen und vernetzte Beziehungen viel komplizierter, doch tut das für die hier verfolgte Fragestellung nichts zur Sache.) Was aber ist die höchste Instanz? Kann sie überhaupt materiell in den Lebewesen gefasst werden? Es stellte sich dann sofort die Frage, was diese Instanz wiederum in Aktion versetzt.
Anwendung: Wie entstanden die fallsgestaltigen Strukturen? Man könnte sich in der anstehenden Frage nach dem Übergang in die Bedingungen und Strukturen nach dem Fall folgendes denken: Die genetischen Grundlagen (die Bausteine als solche) wurden im Gefolge des Sündenfalls nicht geändert, die Instanz aber, die ihren Zusammenbau regelt, reagiert auf die veränderten Lebensbedingungen nach dem Fall. Mit demselben „Baumaterial“, also auf derselben genetischen Grundlage, werden verschiedenartige „Gebilde“ errichtet. Die höchste Instanz also, die die individuelle Entwicklung, den Bau und die Funktionsweisen der Lebensäußerungen regelt, könnte eine Wandlung erfahren haben, so dass unter den neuen Bedingungen nach dem Sündenfall bei vielen Lebewesen neue – fallsgestaltige – Strukturen zum Vorschein kommen.
Dies ist damit vergleichbar, dass man aus demselben Baumaterial ein Rathaus, eine Kirche oder ein Gefängnis bauen kann.
Modell aus der heutigen Biologie. Für diese Vorstellung gibt es ein interessantes Modell aus der heutigen, uns bekannten Biologie: die fremddienliche Zweckmäßigkeit, wie sie in bestechendster Form bei Pflanzengallen zu beobachten ist. Gallen sind spezifisch geformte Gebilde, die vor allem auf Blattoberflächen durch Einwirkung fremder Stoffe (von Bakterien, Pilzen oder Tieren) entstehen. So bildet beispielsweise die Rose nach einem Stich und der Eiablage der Rosengallwespe sog. „Rösenäpfel“ (Abb. 134), büschelige Gebilde, in deren Innern sich einige Kammern für die darin sich entwickelnden Larven befinden (Abb. 135). Es gibt eine reiche Formenvielfalt unter den Gallen. Manche gleichen spitze Hörnern, andere länglichen Zwiebeln, kugelrunden Murmeln, flachen Sonnenhüten, goldglänzenden Münzen oder Knöpfen (Abb. 136). Wichtig ist: Es werden Formen gebildet, die die Wirtspflanze sonst nicht erzeugt. Am gleichen Blatt können sogar verschiedene „Galltiere“ Gallen völlig unterschiedlicher Gestalt hervorrufen. Die neuen Entwicklungswege werden oft mit äußerster Präzision beschritten. Der Stoffwechsel wird zugunsten der Produktion bestimmter Inhaltsstoffe (z. B. Gerbstoffe) umgestellt, manche Wege werden intensiviert, andere verschlossen oder Seitenwege eingeschaltet.

Abb. 134: Ein Rosenapfel (Galle), der von der 3 mm langen Gemeinen Rosengallwespe hervorgerufen wird.
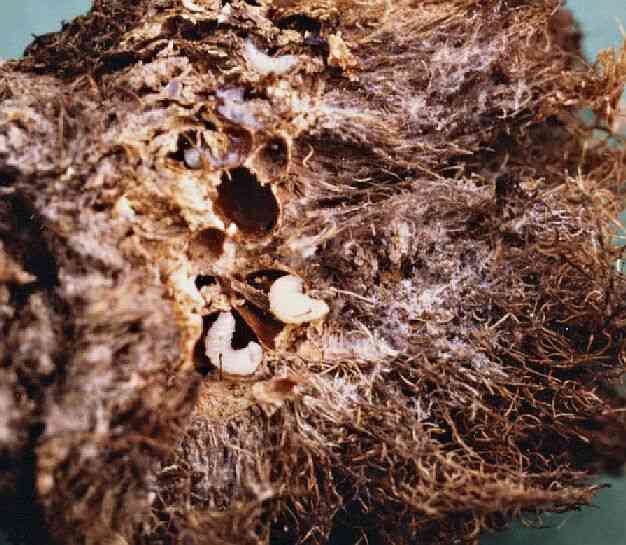
Abb. 135: Der Blick in das Innere eines Rosenapfels offenbart ein Röhrensystem, in dem Larven leben.

Abb. 136: Seidenknopfgallen an der Unterseite eines Eichenblattes, hervorgerufen durch eine Gallwespe
Als Auslöser für die Gallbildung dient den verschiedenen Schmarotzerarten ein Wuchsstoff. Erbsubstanz (DNS) wird jedoch nicht übertragen. I. a. sind die Gallen in komplizierter Weise den Bedürfnissen des Gastes angepasst. Dazu gehören ein passender Hohlraum, ein widerstandsfähiges Gehäuse, zartwandige, der Ernährung dienende Zellen im Innern der Gallen, die Erzeugung bitterer Stoffe, die Vögel oder Raupen vom Fressen abhalten, sowie z. T. die Ausbildung einer Trennschicht, die das Öffnen der Galle ermöglicht, sobald die Insassen zum Ausschwärmen alt genug sind (vgl. Abb. 137).

Abb. 137: Ein fertiges Gallinsekt in einer aufgeschnittenen Eichengalle.
Was geschieht hier? Unter dem Einfluss auslösender Substanzen wird das Baumaterial der Wirtspflanze zum Bau artfremder Strukturen verwendet. Die Gallen ähneln in ihrer Form gewöhnlich irgendwelchen normalen Pflanzenstrukturen nicht im entferntesten. Aber die genetische Grundlage der Pflanzen ist nicht verändert worden. Es werden keine Gene in die Pflanzen injiziert, es erfolgt keine Gentransplantation. Die Gene und der Zellstoffwechsel geraten offenbar unter „fremde Herrschaft“ und werden entsprechend genutzt.
Diesen Beispielen entsprechend könnte man sich denken, dass die geschaffenen Organismen durch den Sündenfall unter eine neue „Herrschaft“, unter eine Art Anpassungstrieb an die Bedingungen „dieses Äons“ gerieten und dadurch ihre Lebensweise änderten. Dieser „Herrschaftswechsel“ muss synchron und bei den verschiedenen Arten aufeinander abgestimmt erfolgt sein, so dass ein nahtloser Übergang in die Ökologie nach dem Sündenfall möglich war. Wichtig ist bei diesem Lösungsversuch, dass die Identität der Arten und Individuen gewahrt bleibt. Zugleich wird so verstehbar, dass das neue komplizierte ökologische Gefüge in seiner Fallsgestaltigkeit koordiniert „zusammengesetzt“ wurde.
Zur Verdeutlichung: Um die Lebensäußerungen der Organismen verstehen zu können, ist das Postulat [= Forderung] einer steuernden Größe sinnvoll. Man könnte es die „Steuerinstanz“ oder „steuerndes Agens“ eines Organismus nennen. Es ist nicht möglich, auf den untergeordneten Ebenen die Organismen zu verstehen. Die „steuernde Instanz“ (die gestaltende Kraft) kann zwar nicht vorgezeigt, jedoch nur an ihren Wirkungen erkannt werden.
Zur Vermeidung eines möglichen Missverständnisses soll noch erwähnt werden, dass Gallenbildung selber nicht als fallsgestaltiges Phänomen zu werten ist, sondern eine Symbiose [= Zusammenleben ohne einseitige Schädigung] darstellt. Das Beispiel soll nur als Illustration dienen, wie sich unter dem Einfluss einer fremden „Größe“ die Gestalt wandeln kann.
Im Rahmen dieses Lösungsversuchs muss nicht angenommen werden, Gott habe (unmittelbar oder latent) fallsgestaltige Anlagen erschaffen.
1.5 Physikalische Rahmenbedingungen
Ein ursprünglich wesensmäßig anderes Ökosystem bedurfte auch anderer physikalischer Rahmenbedingungen als der heutigen. Ohne Tod konnte es auch keine Todesgefahren gegeben haben. Was war z. B. anders, als das Gebären noch nicht schmerzvoll war? Man könnte sich denken, dass die Eigenschaften der Materie von den heutigen Eigenschaften verschieden waren, sodass die anatomischen Merkmale sich nicht schmerzhaft und gefahrvoll auswirkten. Denkbar wäre auch, dass die körperliche Konstitution anders war, so dass die Schmerzen und Gefahren ausgeschlossen waren.
Der Auferstehungsleib Jesu. Eine Gedankenhilfe dafür, dass eine andere Leiblichkeit als die uns bekannte möglich ist, bieten die neutestamentlichen Berichte von der Auferstehung Jesu. Einerseits wird in diesen Schilderungen großen Wert darauf gelegt, dass Jesus leiblich auferstand (er konnte angefasst werden, er aß etwas), andererseits wird bezeugt, dass Jesus durch geschlossene Türen gehen konnte. Wenn wir das versuchen würden, gäbe es mindestens blaue Flecken. Jesu Auferstehungsleiblichkeit war offenbar eine andere, wie auch der Auferstehungsleib der Gläubigen von anderer Qualität sein wird (vgl. 1. Kor 15,42+43). So kann man sich denken, dass die Leiblichkeit des Menschen, die noch nicht durch die Sünde betroffen war, von der heutigen Beschaffenheit verschieden war. Dabei kann jedoch nichts Sicheres darüber gesagt werden, inwiefern die ursprüngliche Leiblichkeit vor dem Sündenfall der künftigen Auferstehungsleiblichkeit gleicht.
1.6 Die zwei Gesichter der Schöpfung – Widerspiegelung des Menschen
Weshalb ist die ganze Schöpfung in den Fall des Menschen mit hineingerissen worden? Eine letztgültige Antwort auf diese Frage soll nicht gegeben werden und ist wohl auch nicht möglich. Römer 8,19ff. hebt diesen Zusammenhang nur hervor, ohne ihn näher zu begründen. Im Artikel „Biblische Aussagen zur Existenzweise der Lebewesen“ (https://genesis-net.de/s/0-5/2-2/ ) wurde bereits auf die schöpfungsgemäße Sonderstellung des Menschen hingewiesen. Wenn das wichtigste Schöpfungswerk „verdorben“ ist, ist alles verdorben, was mit ihm verbunden ist – eben die ganze Schöpfung.
Aus der Verbundenheit zwischen der außermenschlichen Schöpfung und dem Menschen folgt: Die Schöpfung ist eine Widerspiegelung des Menschen und seiner Situation. Die Schöpfung kann sich nicht selber aus der Knechtschaft der Vergänglichkeit befreien, sondern wartet auf den Befreier. Genauso ist es um den Menschen bestellt. Er ist nicht aufgrund eigener Möglichkeiten in der Lage, die Lebensverhältnisse grundlegend zu wandeln, auch er ist von der Vergänglichkeit geknechtet. Alle durchaus notwendigen und richtigen Bemühungen um Verbesserungen der menschlichen Existenz stehen letztlich unter dieser Feststellung. Sie soll nicht zur Resignation führen, sondern zu einem Realismus. Denn in Römer 8,19-22 wird nicht nur eine ernüchternde Diagnose gegeben, sondern auf eine Therapiemöglichkeit hingewiesen. Gott selber, der Unterwerfer, wird auch befreien. Es kann also nur darum gehen, sich dem Schöpfer und seiner Führung anzuvertrauen. Darauf soll uns das „verkehrte“ Gesicht der Schöpfung hinweisen: dass wir ohne Gott auf dem verkehrten Weg sind und seine Befreiung benötigen. Warum seufzt die ganze Schöpfung? „Nicht freiwillig“, sondern „um des willen, der ihre Unterwerfung bewirkt hat“. Damit sind wir gemeint.
Der Zugang zum Schöpfer ist offen, weil Jesus Christus durch sein Sterben und seine Auferstehung diesen Weg eröffnet hat. Jesu Auferstehung bedeutet die Überwindung des Todes und damit der Knechtschaft der Vergänglichkeit. Die Auferstehung ist denen verheißen, die ihm nachfolgen.
1.7 Zusammenfassung
Nach dem Zeugnis der Bibel ist die Welt gefallene Schöpfung und die Daseinsweise der Lebewesen nicht einfach das Ergebnis natürlicher Prozesse. Es wird ein wesensmäßiger Unterschied zwischen der Ursprungswelt bzw. der zukünftigen Schöpfung und der Jetztgestalt der Schöpfung bezeugt. Ökologische Stabilität ist unter den Bedingungen der Jetztzeit („dieses Äons“) nur mit dem Beiklang des Seufzens (Rom 8,19ff.) möglich. Die Frage nach dem Übergang von einer ursprünglichen in die heutige Ökologie ist auf der biologischen Ebene nicht zu beantworten, da es keine entsprechende Erfahrungen aus dem heutigen, uns zugänglichen Bereich gibt. Ein allmählicher Übergang von der ursprünglichen in die heutige Lebensweise der Organismen kommt in der Regel nicht in Frage; es ist meist nur ein abrupter Umbruch denkbar, der die Bedingungen des jetzigen Daseins umfassend ändert. Aus der heutigen Biologie können vage Modelle für diesen Umbruch entnommen werden. Nach einer theologisch befriedigenden Modellvorstellung gerieten die Lebewesen im Gefolge des Sündenfalls unter „Steuerungsinstanzen“, die dem Ursprungsdasein „fremd“, dem jetzigen Dasein ausdrucksgemäß sind. Durch diesen Deutungsansatz sind tiefgreifende Organisationsänderungen und Änderungen der Lebewesen verstehbar, ohne die Notwendigkeit einer zusätzlichen Schöpfung und ohne die Annahme, dass den Lebewesen schon ursprünglich, in der „sehr guten“ Schöpfung bereits genetische Grundlagen für eine andere Lebensweise „eingebaut“ waren.
1.8 Literatur
Junker R (1994) Leben durch Sterben? Schöpfung, Heilsgeschichte und Evolution. Neuhausen-Stuttgart.
Junker R (2001) Sündenfall und Biologie. Schönheit und Schrecken der Schöpfung. Neuhausen-Stuttgart.
Aktualisiert am 07.01.2024 (B. Scholl); © beim Autor: Reinhard Junker, 16.04.2004