Lückenhafte Fossilüberlieferung
Benjamin Scholl (Stand 23.01.2026)
Es ist ein weitverbreitetes Missverständnis, dass die Fossilüberlieferung prinzipiell die Evolutionstheorie nach Charles Darwin bestätigen würde. Im Gegenteil: Wenn heutige evolutionäre Modelle recht hätten, wäre die Fossilüberlieferung in Bezug auf die erwarteten Arten zu 99 % bis sogar 99,997 % unvollständig! Schöpfungswissenschaftler gehen auf der Basis geschaffener Grundtypen (entspricht häufig der Familienebene) ebenfalls von einer lückenhaften Fossilüberlieferung aus. Der folgende Artikel zeigt aber, dass sie keine so große Unvollständigkeit annehmen müssen wie Evolutionsbiologen – wenn auch in anderer Hinsicht und aus anderen Gründen.

Einleitung
Charles Darwin, der Begründer der klassischen Evolutionstheorie und Vordenker der heutigen Evolutionstheorien, war sich der Tatsache bewusst, dass in der Fossilüberlieferung zahlreiche Zwischenformen zwischen heute lebenden und fossilen Taxa fehlen, die man auf Grundlage der Evolutionstheorie erwarten würde (vgl. Kotulla 2020, 3f). Konkret schrieb Darwin (1872/2002, 189): „Wenn Arten aus anderen Arten durch unmerklich kleine Abstufungen entstanden sind, warum sehen wir nicht überall unzählige Übergangsformen? Warum bietet nicht die ganze Natur ein Gewirr von Formen dar, statt dass die Arten, wie sie sich uns zeigen, wohl begrenzt sind?“ (S. 189).
An anderer Stelle fragt Darwin (S. 357): „Woher kommt es dann, dass nicht jede geologische Formation und jede Gesteinsschicht voll von solchen Zwischenformen ist? Die Geologie enthüllt uns sicherlich keine solche fein abgestufte Organismenreihe; und dies ist vielleicht die handgreiflichste gewichtigste Einrede, die man meiner Theorie entgegenhalten kann.“
Um seine Theorie dennoch aufrecht zu erhalten, nahm Darwin (1872, 365) schließlich bei der spekulativen Hilfshypothese Zuflucht, dass die uns bekannte Fossilüberlieferung eben sehr lückenhaft sei (vgl. Junker 2014). Dies könnte einerseits daran liegen, dass Fossilien einfach nur unter sehr speziellen Umständen entstanden sind, und es daher nur von wenigen Arten Fossilien gäbe. Andererseits könnte es auch einfach sein, dass all die zahlreichen „Übergangsformen“, welche Darwin postuliert hatte, bisher noch nicht gefunden worden sind, obwohl sie im Gestein als Fossilien vorhanden sind.
Und so verweisen Evolutionsbiologen bis heute häufig auf diese mutmaßlich extreme Lückenhaftigkeit der Fossilüberlieferung. Schließlich werden ca. 600 Millionen Jahre Evolutionsgeschichte von vielzelligen Lebewesen und deren Vorgeschichte sowie schier unzählige Übergangsformen vorausgesetzt. Somit lautet eine häufige Erklärung der Evolutionsbiologen für die „fehlenden Übergangsglieder“ in der Fossilüberlieferung: Der Prozess des Fossilisierens sei einfach ein unglaublich seltenes Ereignis – so könne man die in der Regel diskontinuierliche Fossilüberlieferung mit abrupt auftauchenden Taxa (Gruppen von Lebewesen) erklären (vgl. Bechly 2022).
Wie lückenhaft ist die Fossiüberlieferung aus der Evolutionsperspektive?
Um diese Frage zu beantworten, werden zwei Zahlen benötigt: 1. Die ungefähre Anzahl an heute bekannten fossilen Arten. Und 2. die Anzahl von Arten, die laut Vorstellungen der Evolutionsbiologen im Laufe der Erdgeschichte gelebt haben sollen.
Die zweite Zahl evolutionär erwartbarer Arten muss deutlich über der heutigen Anzahl von Lebewesen liegen, da jede systematische Ebene von heute lebenden und fossil bekannten Arten schrittweise (graduell) durch Übergangsformen miteinander verbunden sein muss: Dies gilt für alle bekannten Reiche, Stämme, Klassen, Ordnungen, Familien, Gattungen und Arten – sowie für alle zwischen ihnen bestehenden Lücken (mindestens auf genetischer Ebene).
Der Evolutionsbiologe Walter Kleesattel (2001, 22) meint: „Man kennt weniger als 200 000 ausgestorbene Arten von einer Gesamtzahl, die wenigstens 10 Millionen, vermutlich aber ein Vielfaches davon, umfasst hat.“
Alroy (2002, 3706) schätzt, dass man zum damaligen Zeitpunkt ca. 280.000 fossile Arten von Wirbellosen entdeckt hat: „1970 waren etwa 192.000 fossile Arten von Wirbellosen bekannt, und jedes Jahr werden mindestens 3.000 weitere Arten benannt.“ Weiterhin nennt er 3.350[1] valide nordamerikanische fossile Säugetierarten (ebd.).
Geht man davon aus, dass Wirbellose den Großteil der Tiere darstellen, könnte man ableiten, dass heute grob geschätzt ca. 350.000 fossile Spezies von Tieren bekannt sind (so auch Bechly 2022)[2]. Davon sind aber nicht alle ausgestorben, da ca. 6 % der fossil bekannten Arten heute immer noch leben (laut Scholl 2024, Anhang 3 Tab. 1). Im Folgenden rechnen wir der Einfachheit halber mit 350.000 fossil bekannten Arten.
Der weltberühmte Evolutionsbiologe George Gaylord Simpson schätzte die Anzahl der Spezies, die seit Beginn der Evolution diese Erde bevölkert haben sollen, auf 500 Millionen (Spanne von 50 bis 4000 Millionen) – und Cailleux ging von 17 bis 860 Millionen aus (nach Teichert 1956, 967). Davon wurde aber die überwältigende Mehrheit bis heute nicht gefunden, sodass fossil nicht überlieferte Lebensräume in großer Zahl postuliert werden müssen, also Lebensräume in der Erdvergangenheit, aus denen keine Fossilien überliefert worden sind.
Legt man als mittleren Schätzwert ca. 500 Millionen postulierte Spezies in der Evolutionsgeschichte insgesamt und ca. 350.000 fossil bekannte Arten zugrunde, würde dies bedeuten, dass immer noch fast 500 Millionen Spezies seit Beginn ihres evolutionären Auftretens in fossil nicht überlieferten Lebensräumen existiert haben müssten. Dies beträfe über 99,93 % aller fossilen Arten. Ähnlich geht auch Kleesattel (2001, 149) aufgrund der evolutionären Perspektive davon aus, dass 99,9 % aller Arten „mit ziemlicher Sicherheit“ wieder ausgestorben sind. Auch Schweitzer et al. (2021, 43) geben „weit über 99 % aller Arten“ an; bei Barnosky et al. (2011, 51) sind es nur rund 99 % der Arten.
Ähnlich heißt es auch auf spektrum.de in einem populären Artikel: „Im Lauf der Erdgeschichte gab es schätzungsweise vier Milliarden Pflanzen- und Tierarten. Davon sind 99 Prozent verschwunden. Viele von ihnen haben jedoch Spuren hinterlassen“ (Hemmer & Meßner 2025). Der Hinweis, dass „viele“ der 4 Milliarden Jahren Spuren – also Fossilien – hinterlassen haben, ist hier aber völlig unangemessen: Bei 4 Milliarden Spezies – das ist auch die Obergrenze der Schätzung von Simpson (s. o.) – würden ca. 99,99125 % aller fossilen Arten fehlen.
Und Barker (2023, 10) meint gar: „Seit den Anfängen des Lebens haben etwa 5 Milliarden Arten die Erde bewohnt. Mehr als neunundneunzig Prozent von ihnen sind heute ausgestorben.“ Dementsprechend würden sogar 99,993 % der fossilen Arten fehlen.
Zrzavý et al. (2013, 365–367) nennen in ihrem Standardlehrbuch für Evolution heute ca. 2 Millionen beschriebene Arten und schätzen den heutigen Bestand auf 4 bis 6 Millionen Arten (bzw. „ein paar wenige Millionen“) – dabei begründen sie auch, warum die weitverbreitete These, dass es 30 Millionen heutige Arten geben könnte, die Datenlage wohl deutlich überschätzt. Dann schreiben die Autoren (S. 367): „Die geschätzte Gesamtzahl der Arten, die unsere Erde je bewohnt haben, ist noch um drei Größenordnungen höher. Das heißt, dass sich während der gesamten Erdgeschichte ca. zehn Milliarden Speziationen abgespielt haben, wobei jedes tausendste Speziationsereignis [Artbildungsereignis] eine Evolutionslinie hinterlassen hat, deren Nachkommen bis heute überlebt haben.“ Bei ca. 350.000 fossilen Arten fehlen dann also 99,9965 % aller fossilen Spezies aus evolutionärer Perspektive!
Kurzum: In Bezug auf die pure Artenzahl fehlen laut anerkannten Evolutionsbiologen zwischen 99 % und 99,9965 % aller evolutionär postulierten Arten in der fossilen Überlieferung. Es gibt also eine gewaltige Diskrepanz zwischen realen Daten (Fossilien) und Deutungen (evolutionären Stammbaumhypothesen).
Doch selbst die fossilen Arten, die man gefunden hat, sind in vielen Fällen nicht die Übergangsformen, die man sich erhofft hatte, um die heute existenten Arten damit in einem evolutionären Stammbaum verknüpfen zu können.
Eine weitere Unvollständigkeit der Fossilüberlieferung aus evolutionärer Perspektive ergibt sich aus den Lücken zwischen den evolutionär aufgrund molekulargenetisch errechneten Erscheinungszeitpunkten von Taxa (Gruppen wie Arten, Gattungen etc.) einerseits und den tatsächlichen Zeitpunkten des erstmaligen fossilen Nachweises andererseits. Man spricht hier von Geisterlinien (vgl. Scholl 2024 Anhang 4 sowie Anhang 1 Tab. 2 für die Fische).[3]
Weitere Zitate von Evolutionsbiologen zur Frage der Unvollständigkeit der Fossilüberlieferung befinden sich im Anhang.
Wie unvollständig ist die Fossilüberlieferung aus Schöpfungsperspektive?
Schöpfungswissenschaftler gehen nicht von einzelnen Arten (Biospezies) aus, die in einer schrittweise sich evolvierenden Kette von Lebewesen „von der Amöbe bis zu Goethe“ entstanden sein sollen. Für sie ist die relevante Einheit nicht die Art (Biospezies), sondern die von Gott erschaffenen Schöpfungsarten (Grundtypen), die sich aufgrund ihres ursprünglich vielfältigen vorprogrammierten Erbguts in verschiedene Spezies ausdifferenzieren können. Häufig liegen Grundtypen auf dem mittleren systematischen Niveau von Familien, wenn teilweise auch leicht darüber oder darunter (vgl. Kutzelnigg & Scholl 2025). Mittlerweile sind verschiedene Mechanismen bekannt, die schnelle Artbildungsprozesse innerhalb von Grundtypen ermöglichen, wie Mendel‘sche Rekombination und epigenetisch gesteuerte Umlagerung (s. z. B. Crompton 2018 sowie Borger 2022). Solche Prozesse sind nicht auf langsame, schrittweise (graduelle) Selektions-Mutations-Prozesse angewiesen, wie Darwin’sche Evolutionsmodelle. Stattdessen ermöglichen sie sehr schnelle Artbildungsprozesse in wenigen Generationen, sodass eine fossile Überlieferung der Prozesse im Einzelnen oft gar nicht erwartbar ist. Dementsprechend sind Familien – und nicht Arten wie bei der graduellen Evolution – die relevantesten systematischen Einheiten, deren Auftreten in der Fossilüberlieferung aus Schöpfungsperspektive zu bewerten und zu deuten ist.
Schöpfungswissenschaftler wissen nicht, wie viele Grundtypen Gott ursprünglich erschaffen hat. Im biblischen Grundtyp-Modell kann man aufgrund von bspw. Genesis 2,1f, Exodus 20,11 und Hebräer 4,3f aber davon ausgehen, dass nach der Schöpfungswoche keine neuen Grundtypen mehr erschaffen wurden.[4] Dementsprechend kann man schlussfolgern, dass sich durch Aussterbeereignisse der Anteil der überlebenden Grundtypen nach der Schöpfung (bzw. seit dem Sündenfall) stetig verringert hat[5] – dies gilt umso mehr, je stärker die geologischen und ökologischen Umwälzungen waren. So ist es sehr wahrscheinlich, dass es gewaltige geologische und ökologische Umwälzungen im Rahmen der Sintflut gab (gemäß Genesis 6–8), die die Grundtypen unter massiven Selektionsdruck gebracht haben. Je weiter Zeit man in der Vergangenheit zurückgeht, umso mehr heute ausgestorbene Grundtypen sind daher in der Fossilüberlieferung zu erwarten. Für Schöpfungswissenschaftler ist somit vor allem die Frage relevant, inwiefern sich heutige Grundtypen als lebende Fossilien in der Fossilüberlieferung ausmachen lassen. Und wo insofern Lücken bestehen, als dass heutige Grundtypen in tieferen geologischen Schichten fehlen.
Ein erster Anhaltspunkt findet sich bei Zrzavý et al. (2013, 416, Abb. 6.19) in einer Abbildung mit fossilen marinen Familien über die geologischen Systeme seit dem Ediacarium (das vor ca. 539 MrJ endete). Bis auf die Mikrofossilien findet sich die gleiche Darstellung hier als Abb. 1. Die Gruppen sind (relativ) subjektiv eingeteilt – von Interesse sind an diesem Punkt vor allem die „Modernen“ Familien (obwohl heute auch noch andere Gruppen existieren). Die Anzahl von grob 650 fossilen „Modernen Familien“ am Ende des Känozoikums (Erdneuzeit, nach der Kreide vor ca. 66 MrJ) fällt in der Retrospektive über das Erdmittelalter (Trias bis Kreide) moderat ab – so dass fast noch 200 moderne Familien bis ins mittlere Karbon im oberen Erdaltertum gefunden werden. Allerdings fällt deren Teil bis ins untere Erdaltertum (Ordovizium und Kambrium) noch stärker ab, so dass nur einige wenige „Moderne“ Familien auch dort gefunden werden. Aus Kurzzeit-Schöpfungsperspektive würde man aber erwarten, dass die Fossilüberlieferung (wenigstens der Tiere) wohl erst nach dem Sündenfall stattgefunden hat (vgl. Genesis 1,30; Römer 8,20f) und dass also auch im Erdaltertum die heute noch lebenden Grundtypen vertreten waren. Daher müsste man deutliche Lücken der Fossilüberlieferung im frühen Erdaltertum und moderate Lücken von heutigen Tierfamilien im Erdmittelalter annehmen.

Abb. 1 Ein Überblick über marine Familien der Tiere („Fauna“) in den geologischen Systemen (nach Bokulich 2021, Fig. 1). Die Einteilung in „Kambrische“, „Paläozische“ und „Moderne“ Familien ist insofern subjektiv, als auch in der Erdneuzeit (Känozoikum) „Paläozoische“ Familien in der Fossilüberlieferung vorkommen – und „Moderne“ Familien im Kambrium. (Eigene Darstellung nach Bokulich 2021, Fig. 1 – diese basiert auf Sepkoski 1984).
Abb. 1 bleibt allerdings nur ein allererster Anhaltspunkt, weil die Daten fast 40 Jahre alt sind und die Einteilung der Familien in die Gruppen sehr unkonkret ist. Daher wurde eine eigene Analyse auf Grundlage der umfassenden Fossilien-Datenbank PBDB (The Paleobiology Database) durchgeführt (s. Scholl 2024, Anhang 3).
Dabei ergibt sich, dass von allen aufgeführten fossilen Familien 5807 ausgestorben, aber 3494 heute noch lebend sind. Das bedeutet: 37,57 % der fossilen Familien existieren heute noch. Diese lassen sich durchschnittlich (!) insgesamt 103,08 bis 110,77 Millionen radiometrische Jahre in der Erdvergangenheit zurückverfolgen – also bis in die Unterkreide.
Geht man vom einem Beginn der Fossilüberlieferung komplexen makroskopischen Lebens vor maximal 600 MrJ aufgrund von präkambrischen Makrofossilien aus der Lantian-Formation (Südchina) aus (vgl. Junker 2014; Narbonne 2011), dann fehlen heutige Familien im Durchschnitt in 82,18 % der Fossilüberlieferung und sind in ca. 17,82 % vorhanden (= 107 MrJ / 600 MrJ).
Heutige Familien, die in der Fossilüberlieferung auftreten, sind also im Durchschnitt 107 MrJ alt, was immerhin ca. 18 % der Fossilüberlieferung makroskopischer Lebensformen entspricht – auch wenn hier jeweils andere Ebenen zugrunde liegen, die einen direkten Vergleich mit den evolutionstheoretischen Erwartungen nicht direkt ermöglichen. Familien als „lebende Fossilien“ aus dem Erdmittelalter sind somit die Regel und nicht die Ausnahme.
In Anbetracht der Collector-Curve (Sammlerkurve), die aufzeigt, wie viele neue fossile Arten pro Jahr gefunden worden sind, ist auch nicht davon auszugehen, dass sich diese Verhältnisse noch um Größenordnungen verschieben werden – immerhin zeichnet sich bereits seit den 1950er-Jahren eine Sättigung ab (vgl. Scholl 2024, Anhang 3), wobei es in manchen Gruppen Ausnahmen gibt (z. B. bei Vogelfamilien in der Kreide). Fossile Familien sind aus Schöpfungsperspektive in fast einem Fünftel der makroskopischen Fossilüberlieferung vorhanden und die Zahl ausgestorbener fossil bekannter Familien ist nicht einmal doppelt so groß wie Anzahl heutiger Familien. Es kann also durchaus sein, dass Gott nicht um Größenordnungen mehr Grundtypen erschaffen hat, als wir heute kennen. Damit ist die Fossilüberlieferung aus Schöpfungsperspektive m. E. nicht so unvollständig wie aus Evolutionsperspektive.
Fazit
Die Fossilüberlieferung ist aus evolutionärer Perspektive, die von einer artenbasierten, graduellen Evolution ausgeht, extrem unvollständig und zwar zu 99 % bis 99,9965 %. Evolutionäre, fossile „Übergangsformen“ fehlen in gewaltigem Ausmaß. Auch machen evolutionäre Stammbaummodelle (bzw. Strato-Cladogramme) die Annahme vielfacher Geisterlinien notwendig. Evolutionäre Deutungen müssen sich somit auf einen winzigen Anteil greifbarer Daten stützen und über den gewaltigen Rest Mutmaßungen treffen.
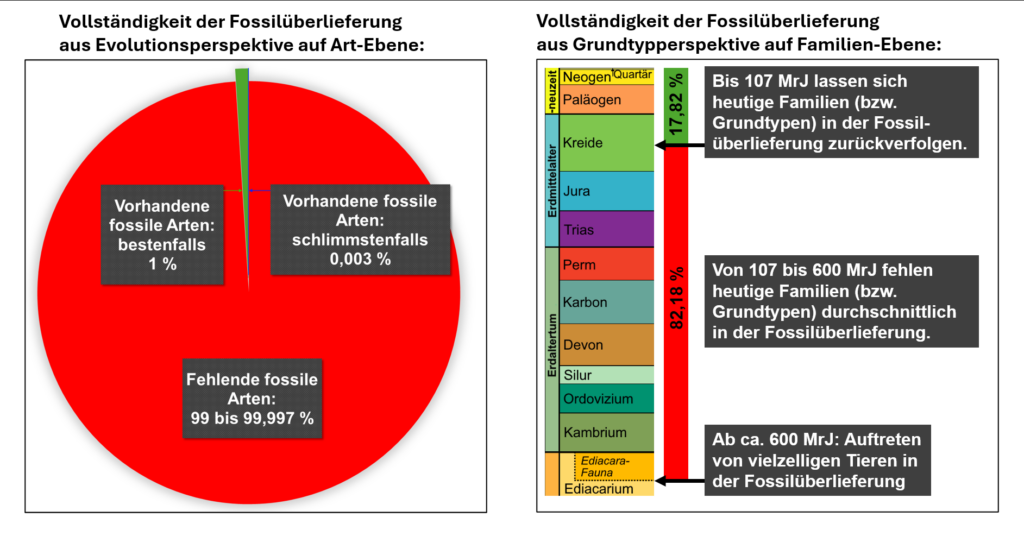
Abb. 2 Links: Unvollständigkeit der Fossilüberlieferung aus Evolutionsperspektive (wegen der fehlenden Übergangsformen) auf Art-Ebene; für das Worst-Case-Szenario (blau) muss man sehr genau ranzoomen, um das noch zu sehen. Rechts: Unvollständigkeit der Fossilüberlieferung aus Grundtypperspektive auf Familien-Ebene. Wichtig: Die Prozentzahlen sind nicht direkt vergleichbar, da sie anderen Sachverhalten entstammen. (Eigene Darstellung)
Aus Schöpfungsperspektive ist die Fossilüberlieferung ebenfalls unvollständig, wenn auch meist in ganz anderer Hinsicht, auf ganz anderer Ebene und aus anderen Gründen. Hier stellt sich die Frage, wo die heutigen Grundtypen – bzw. sich schnell diversifizierender Familien statt Arten in der graduellen Evolution – in der Fossilüberlieferung vor ihrem ersten fossil bekannten Auftauchen waren. Es müssen also ebenfalls Geisterlinien und fossil nicht überlieferte Lebensräume angenommen werden. Diese betragen allerdings „nur“ ca. 82,18 % des geologischen Zeitraums (in radiometrischen Jahren) seit dem ersten fossilen Auftreten des makroskopischen Lebens am Ende des Präkambriums. Schließlich sind lebende Familien im Durchschnitt in 17,82 % der Fossilüberlieferung seitdem nachgewiesen. Die Collector Curve der Vergangenheit spricht dafür, dass sich an diesem Befund auch nicht viel ändern wird. Dennoch besteht weiter Forschungsbedarf zu der Geschwindigkeit der Artaufspaltung von Grundtypen, zu dem systematischen Level von Grundtypen und zu der Frage, wie die Abfolge fossiler Lebensgemeinschaften mit einem biblischen Sintflutmodell in Übereinstimmung zu bringen ist – und warum heutige Familien in 82 % der genannten geologischen Schichten fehlen.
Dennoch bleibt festzuhalten: Was die Fossilüberlieferung angeht, müssen sich schöpfungsgläubige Christen nicht vor evolutionsgläubigen Atheisten verstecken.
Anhang: Zitate-Sammlung bezüglich der aus evolutionärer Perspektive unvollständigen bzw. unerwarteten Fossilüberlieferung
Im Folgenden finden sich Zitate von Evolutionsbiologen, die weitere Aspekte der Lückenhaftigkeit des Fossilberichts aus evolutionärer Perspektive zeigen (kursive Hervorhebungen sind hinzugefügt).
„Trotz dieser Beispiele bleibt es wahr, wie jeder Paläontologe weiß, dass die meisten neuen Arten, Gattungen und Familien und fast alle neuen Kategorien oberhalb der Familien plötzlich in den Aufzeichnungen erscheinen und nicht durch bekannte, allmähliche, vollständig kontinuierliche Übergangssequenzen angeführt werden.“ (George G. Simpson [1953] The Major Features of Evolution. Columbia University Press, S. 360, nach: https://www.talkorigins.org/faqs/quotes/ mine/part1-2.html, aufgerufen am 07.12.2022.) Es empfiehlt sich, dieses Zitat im erweiterten Kontext auf der angegebenen antikreationistischen Website nachzulesen, da es zeigt, dass evolutionäre Modelle zwar theoretische Erklärungsangebote für diesen Befund aufweisen können, der fossile Befund, der im Zitat beschrieben wird, aber real ist. So erklärt Simpson (1953, ebd.) beispielsweise: „Die Chance, dass die Überreste eines Organismus begraben werden, versteinern, im Gestein bis heute erhalten bleiben, dann an der Oberfläche des trockenen Landes freigelegt und von einem Paläontologen gefunden werden, bevor sie zerfallen, ist äußerst gering, praktisch verschwindend gering. Die Entdeckung eines Fossils einer bestimmten Art – unter den Tausenden von Millionen, die die Erde bevölkert haben –, erscheint selbst einem Paläontologen […] fast wie ein Wunder.“ Allerdings kann mit solchen unsystematisch zugänglichen „Wundern“ wohl kaum valide eine umfassende Theorie über die Vergangenheit begründen.
Auch Benton (2009, 121) schlägt als Erklärung für den lückenhaften Fossilbefund vor: „Zum Beispiel korreliert die Aufzeichnung der Vielfalt im Laufe der Zeit eng mit der Gesteinsaufzeichnung […]. Könnte es sein, dass das Muster der Fossilvielfalt einfach eine Aufzeichnung der Gesteinsverfügbarkeit ist? Wenn in einem bestimmten Zeitabschnitt viel Sedimentgestein abgelagert wurde, könnten viele Fossilien gefunden werden.“
Zrzavý et al. (2013, 403f) schreiben: „Eine Gruppe von Paläontologen um Stephen Jay Gould […] und Niles Eldredge […] legte Anfang der 70er-Jahre des 20. Jahrhunderts eine Alternative vor, mit der Begründung, dass fast nirgendwo graduelle Änderungen zu sehen sind. Diese an sich unbestrittene Tatsache erklärte man üblicherweise mit der Unvollständigkeit fossiler Funde: Selbst, wenn die Evolution graduell verlief, würden sowieso nur isolierte Punkte fossil erhalten bleiben. […] Der Mangel an ‚Übergangsgliedern‘ entsteht dadurch, dass die Evolution der Neuheiten nur in kleinen und schwer zu entdeckenden Populationen irgendwo an der Peripherie des Verbreitungsgebiets der ursprünglichen Art stattfindet. Im Regelfall ist das nicht dort, wo wir etwas ausgegraben haben. Die neue Art kann nur durch nachträgliche Migration in das Gebiet ihres Vorfahren gelangen – und erscheint dort dann relativ plötzlich im Fossilbericht“.
Bei Junker (2014; z. T. in dt. Übersetzung) finden sich einige Feststellungen zu diesem Thema, z. B.: „‚Die Fossilienaufzeichnungen enthalten zahlreiche Belege für zwei scheinbar widersprüchliche Muster: Ausbrüche morphologischer Innovationen, die oft mit adaptiven Radiationen innerhalb von Gruppen in Verbindung gebracht werden, und langfristige Stabilität der Morphologie‘ (Davidson & Erwin 2010, 184)“ (Junker 2014, 35; auf S. 11 führt er auch Erwin & Valentine 2013, 157 hierzu an). Das widerspreche aber evolutionstheoretischen Erwartungen (Junker 2014, 11). „‚Wenn jede dieser Hauptgruppen im Fossilbericht auftaucht, umfasst sie einen Großteil, wenn nicht die gesamte morphologische Bandbreite (Disparität), die in der späteren Geschichte der Gruppe zu finden ist. Wenn wir in der Linne‘schen Hierarchie der Stämme, Klassen, Ordnungen usw. nach unten gehen, stellen wir fest, dass alle marinen Stämme (mit Ausnahme der Bryozoen [Moostierchen]) bereits im frühen Kambrium auftraten und es keine Anzeichen für Zwischenformen zwischen den Stämmen gibt (Erwin et al. […]). Die meisten der großen Klassen, die auch im Kambrium vorhanden waren, sind in Bezug auf die gemeinsamen Merkmale, durch die die Klassen definiert werden, seither im Wesentlichen unverändert geblieben’ (Davidson & Erwin 2010, 184)“ (Junker 2014, 35). „Der Befund der frühen und bald maximalen Verschiedenartigkeit gilt nicht nur für die kambrischen Formen, sondern auch für Untergruppen, die erst in jünger datierten geologischen Systemen fossil in Erscheinung treten. Dies hat eine im Jahr 2013 veröffentlichte systematische Studie über die Fossilabfolgen verschiedener Organismengruppen gezeigt (Hughes et al. 2013). Demnach wird der Höhepunkt der Verschiedenartigkeit der untersuchten Gruppen frühzeitig erreicht und nimmt im Laufe der Fossilüberlieferung ab“ (Junker 2014, 12). „‚… es ist klar, dass diese Kladen, sobald sie einmal etabliert waren, Millionen von Jahren überdauerten. Diese biotische Stabilität steht jedoch im Gegensatz zu den sich verändernden Bedingungen des kambrischen Meeresbodens‘ (Erwin & Valentine 2013, 157)“ (Junker 2014, 35). „‚[I]n der Mehrzahl der untersuchten kambrischen und frühpaläozoischen Kladen, im Allgemeinen auf Klassenebene, übersteigt die morphologische Disparität zunächst die taxonomische Vielfalt bei weitem. … Es ist genau das Gegenteil der Erwartung, dass die morphologische Vielfalt im Laufe der Zeit allmählich zunimmt‘ (Erwin & Valentine 2013, 341)” (Junker 2014, 35).
Auch bei Voje et al. (2018) wird deutlich, dass Stasis ein Problem der Fossilüberlieferung ist, das Evolutionsbiologen mit verschiedenen Evolutionsmodellen zu ergründen suchen: „Langfristige phänotypische Stasis wird häufig im Fossilbericht beobachtet […]. Das Verständnis langfristiger morphologischer Stasis (d. h. geringe oder keine Nettoevolution in einer Linie auf makroevolutionären Zeitskalen; Eldredge und Gould 1972; Gould und Eldredge 1977; Gould 2002) bleibt eine große Herausforderung in der Evolutionsbiologie (Bradshaw 1991; Hansen und Houle 2004; Voje 2016)“ (Voje et al. 2018, 509). „Darüber hinaus sagt die populationsgenetische Theorie schnellere Veränderungsraten bei quantitativen Merkmalen voraus als das, was üblicherweise im Fossilbericht beobachtet wird (Lynch 1990; Cheetham et al. 1994) […]. Dennoch scheint Stasis ein üblicher Modus der Evolution im Fossilbericht zu sein (Hunt 2007; Hopkins und Lidgard 2012; Hunt et al. 2015; Voje 2016)“ (ebd.).
Hössjer et al. (2021, 2) fassen das evolutionäre Problem des Fossilbefundes folgendermaßen zusammen und nennen zahlreiche Beispiele (vgl. noch ausführlicher bei Bechly 2024): „Der Fossilnachweis wird häufig so interpretiert, dass es lange Perioden der Stasis gibt (Voje et al., 2018), die von abrupteren Veränderungen und ‚explosiven’ Ursprüngen unterbrochen werden (Bechly und Meyer, 2017). Zu diesen Veränderungen gehören zum Beispiel die Evolution des Lebens (Bell et al., 2015), die Photosynthese (Hecht, 2013), die Vielzelligkeit und die ‚Avalon Explosion’ (Shen et al., 2008), Tierkörperpläne und die ‚Kambrische Explosion’ (Erwin und Valentine, 2013), komplexe Augen (Paterson et al., 2011), Kiefer und Zähne von Wirbeltieren (Fraser et al., 2010), Terrestrialisierung (z. B. bei Gefäßpflanzen, Arthropoden und Tetrapoden) (Bateman et al., 1998), die Metamorphose von Insekten (Labandeira, 2011), das Flugverhalten und die Federn von Tieren (Wu et al., 2018; Yang et al., 2019), die Fortpflanzungssysteme, einschließlich der Blütenstände von Angiospermen, Eier von Amnioten und die Plazenta von Säugetieren (Chuong, 2013; Doyle, 2012; Roberts et al, 2016; Sauquet, 2017; Specht und Bartlett, 2009), Echolokation bei Walen (Churchill et al., 2016; Park et al., 2016) und Fledermäusen (Simmons et al., 2008) […]. Basierend auf der radiometrischen Datierung der verfügbaren Zeitfenster im Fossilbericht wird angenommen, dass diese genetischen Veränderungen auf einer makroevolutionären Zeitskala sehr schnell stattgefunden haben.“
Unter Evolutionsbiologen ist die Diskussion um die Vollständigkeit der Fossilüberlieferung trotz (oder vielleicht gerade wegen) ganz verschiedener Ansätze[6] zu deren Ermittlung in der Fachwelt nicht annähernd entschieden, wie auch folgende Zitate zeigen:
- Benton (2009, 117) schreibt in einem dezidiert antikreationistischen Artikel: „Zeigt uns die Fossilüberlieferung 50 % oder 90 % oder 99,99999 % der ausgestorbenen Arten? Das kann natürlich nie festgestellt werden.“ Interessanterweise nennt er die Option ca. 1 % bis 0,0035 % gar nicht, die aus evolutionärer Perspektive wohl am ehesten zu erwarten wäre (s. o.).
- „Der Unterschied zwischen dem potenziellen [maximal zu entdeckenden] Fossilbestand und der Realität kann sehr groß […]“ sein (Benton et al. 2011, 64).
- „Es wurden zwei gegensätzliche Standpunkte vertreten: Entweder ist der Fossilnachweis gut genug […] oder nicht gut genug […], um die wichtigsten Muster der globalen Diversifizierung im Laufe der Zeit aufzuzeigen. Eine Auflösung zwischen diesen gegensätzlichen Standpunkten scheint nicht naheliegend“ (Benton et al. 2011, 63).
- „Jeder Zeitabschnitt, jede geografische Region und jede Clade [Gruppe] ist anders beprobt, so dass eine globale Antwort wahrscheinlich nie gefunden werden kann“ (Benton et al. 2011, 90f).
- „Eine eindeutige Antwort auf die Frage, ob der Fossilienbestand durch Makroevolution oder Megabias [Verzerrung] bestimmt wird, wird es aufgrund der zeitlichen, geografischen und taxonomischen Varianz in den Daten wahrscheinlich nie geben“ (Benton et al. 2011, 63).
Literatur
Alroy J (2002, 3706) How many named species are valid? PNAS 99, 3706–3711, www.pnas.org/ cgi/doi/10.1073/pnas.062691099.
Barker C (2023) Das ultimative Dinosaurier Lexikon. Übersetzt von Ehrhardt K. Ravensburger Verlag. Ravensburger.
Bechyl G (2022) Fact Check: Hawaiian Silverswords Fail the Species Pair Challenge. Evolution News, veröffentlicht am 03.05.2022, https://evolutionnews.org/2022/05/fact-check-hawaiian-silverswords-fail-the-species-pair-challenge/.
Bechyl G (2024) Fossil Friday: Discontinuities in the Fossil Record — A Problem for Neo-Darwinism, vom 10.05.2024, https://evolutionnews.org/2024/05/fossil-friday-discontinuities-in-the-fossil-record-a-problem-for-neo-darwinism/.
Benton M (2009) The completeness of the fossil record. Significance 6, 117–121, https://doi.org/10.1111/ j.1740-9713.2009.00374.x.
Benton M, Dunhill A, Lloyd G & Marx F (2011) Assessing the quality of the fossil record: Insights from vertebrates. Geol. Soc. Spec. Publ.358, 63–94, https://doi.org/10.1144/SP358.6.
Binder H (2018) Neues von Bernstein-Inklusen. Unerwartete Einsichten zur Geschichte von Gliederfüßern. Stud. Integr. J. 25, 96–100.
Borger P (2022) Epigenetik und programmierte Anpassungen. Genesisnet News vom 12.04.2022, https://genesis-net.de/n/300-0/.
Crompton N (2018) Mendel’sche Artbildung und die Entstehung der Arten. W+W Special Paper B-19-3, https://www.wort-und-wissen.org/artikel/mendelsche-artbildung-und-die-entstehung-der-arten/.
Darwin C (1872; dt. Übersetzung 2002) Über die Entstehung der Arten durch natürliche Zuchtwahl oder die Erhaltung der begünstigten Rassen im Kampfe um’s Dasein. Nach der letzten englischen Auflage wiederholt durchgesehen von J. Victor Carus. Parkland Verlag.
Hemmer R & Meßner D (2025) Kleine Geschichte über die Entdeckung der Massenaussterben, vom 08.01.2025, https://www.spektrum.de/kolumne/hemmer-und-messner-kleine-geschichte-der-entdeckung-des-aussterbens/2211857.
Hössjer O, Bechly G & Gauger A (2021) On the waiting time until coordinated mutations get fixed in regulatory sequences. J. Theor. Biol. 524, 110657, https://doi.org/10.1016/j.jtbi.2021.110657.
Junker R (2014) Zur neueren Diskussion über die kambrische Explosion. W+W Special Paper B-14-1, https://www.wort-und-wissen.org/wp-content/uploads/b-14-1_kambrische_explosion.pdf.
Kleesattel W (2001) Die Welt der Lebenden Fossilien. Eine Reise in die Urzeit. Wissenschaftliche Buchgesellschaft, Darmstadt.
Kutzelnigg H & Scholl B (2025) Zusammenstellung vorläufiger Grundtypen (Auswahl). W+W-Onlineartikel vom 25.02.2025, https://www.wort-und-wissen.org/artikel/grundtypentabelle/.
Kotulla M (2020) Gültigkeit und Grenzen geologischer Zeitbestimmung. Online-Loseblattsammlung 2-22, Stand: 10/2020. The Art of Charles Darwin: Unermessliche Zeit und unvollständige Fossilüberlieferung (v2010), https://www.wort-und-wissen.org/publikationen/geologie-loseblattsammlung/.
Lidgard S & Love AC (2018) Rethinking Living Fossils. BioScience 68, 760–770, https://doi.org/10.1093/biosci/biy084.
Narbonne G (2011) When life got big. Nature 470, 339–340, https://doi.org/10.1038/470339a.
Schweitzer MH, Schroeter ER & Czajka CD (2021) Dinosaurs. How We Know What We Know. CRC Press. Boca Raton & Oxon.
Suling M (2021) Kretazische Insekten im Gestein und als Einschlüs-se. W+W Special Paper B-21-2, https://www.wort-und-wissen.org/wp-content/uploads/b-21-2_kretazische-insekten.pdf.
Teichert C (1956) How Many Fossil Species? J. Paleontol. 30, 967–969, https://www.jstor.org/stable/1300432.
Voje KL, Starrfelt J & Liow LH (2018) Model Adequacy and Microevolutionary Explanations for Stasis in the Fossil Record. Am. Nat. 191, 509–523, doi: 10.1086/696265.
Zrzavý J, Storch D & Mihulka S (2013) Evolution. Ein Lese-Lehrbuch. Hrsg. der deutschen Ausgabe: Burda H & Begall S. 5. Aufl. Spektrum Akademischer Verlag Heidelberg.
Anmerkungen
[1] Alroy (ebd.) geht davon aus, dass ca. 24–31 % der aktuell benannten fossilen nordamerikanischen Säugetierarten nicht valide sind – sonst wäre die Zahl noch höher.
[2] Bechly (2022a) schreibt: „Unter den 350.000 beschriebenen fossilen Arten können wir zahlreiche abrupte Ursprünge neuer Körperformen innerhalb eines Zeitfensters von 5–10 Millionen Jahren feststellen. Unter den schätzungsweise 8,7 Millionen rezenten [lebenden] Arten finden wir keine derartigen Unterschiede im Körperbau bei Artenpaaren, die sich nach Untersuchungen der molekularen Uhr in einem ähnlichen Zeitrahmen entwickelt haben. Dies widerspricht den Erwartungen aus einer darwinistischen Perspektive.“
[3] Bei Fischen finden sich evolutionäre Geisterlinien zuhauf (s. Scholl 2024, z. B. Anhang 3). Auch bei anderen Gruppen finden sich häufig evolutionär postulierte Geisterlinien im Bereich von meist ein bis zwei Dutzend MrJ. Vgl. z. B. die Archosaurier bei Benton et al. (2011, Fig. 8). Benton et al. (2011, 83) schreiben zudem: „Der Fossilnachweis mesozoischer Vögel enthält sicherlich sehr viele Geisterbereiche (55 Geisterbereiche auf [stratigrafischer] Stufenebene und nur 29 Aufzeichnungen auf Stufenebene)“.
[4] „So wurden vollendet der Himmel und die Erde und all ihr Heer. Und Gott hatte am siebten Tag sein Werk vollendet, das er gemacht hatte“ (Genesis 2,1f); und: „Denn in sechs Tagen hat der HERR den Himmel und die Erde gemacht, das Meer und alles, was in ihnen ist, und er ruhte am siebten Tag“ (Exodus 20,11; je Elberfelder CSV Übersetzung). „Nun waren ja die Werke von Anbeginn der Welt gemacht; denn so hat er an einer andern Stelle gesprochen vom siebenten Tag: ‚Und Gott ruhte am siebenten Tag von allen seinen Werken.‘“ (Hebräer 4,3c–4)
[5] An dieser Stelle wird auf eine Diskussion zu der Fragestellung, ob Grundtypen auch innerhalb oder gar vor der Schöpfungswoche schon hätten aussterben können, sowie auf die Frage der Dauer der Schöpfungswoche verzichtet, sondern es wird vom Tod der Tierwelt nach dem Sündenfall (Genesis 3) ausgegangen. Vgl. dazu z. B. Stephan (2015, 21–37) sowie ausführlich Junker (2017).
[6] Ein Beispiel: Binder (2018) hat auf der Grundlage einer Studie mit heutigen Gliederfüßern darauf hingewiesen, dass im Fall von Bernstein die direkt in der Nähe lebenden Gliederfüßer (v. a. Insekten und Spinnen) eine gute Chance zur Fossilisation in Bernstein haben. Allerdings scheint sich nicht die komplette Artvielfalt des Ökosystems im Bernstein zu erhalten. Vgl. detaillierter zum Erhalt von Insekten in Sediment- und Bernsteineinschlüssen: Suling (2021).
Dieser Artikel ist eine leicht aktualisierter Textauszug aus: Scholl B (2024) Die ältesten fossilen Fische: Große Vielfalt seit Beginn der fossilen Überlieferung. W+W-Onlineartikel vom 08.08.2024, https://www.wort-und-wissen.org/artikel/die-aeltesten-fossilen-fische/.