Update: „Darwin Devolves“ (Expertentext über Michael Behes neuestes Buch)
In seinem letzten Buch mit dem Titel „Darwin Devolves“ vertritt der Biochemiker und Verfechter von Intelligent Design (ID), Michael Behe, die These, dass Darwin’sche Evolution „stark überwiegend“ destruktiver Natur ist. Evolution geht langfristig einher mit Verlust bzw. Beschädigung genetischer Information und der dazu gehörenden biologischen Funktionen und könne daher keine wirkliche Innovation in der Biologie bewirken. Dazu hat es viele kritische Stellungnahmen gegeben, auf die Behe wiederholt antwortete. Wessen Argumente sind überzeugender?
Boris Schmidtgall (Artikel „Evolution oder Devolution? Wohin „geht“ die Entwicklung der Arten?“ aus Studium Integrale Journal 30,94–102)
Zusammenfassung
Die Auffassung, dass die gesamte Naturgeschichte ausgehend von den ersten Organismen bis zur heutigen Artenvielfalt eine Höherentwicklung im Verlauf von vier Milliarden Jahren durchlaufen hat, stellt immer noch die vorherrschende Überzeugung bei Naturwissenschaftlern dar. Der Biochemiker Michael Behe stellte diese Denkweise in seinem jüngsten Buch „Darwin Devolves“ in Frage, indem er darlegte, warum die Evolutionsfaktoren Mutation und Selektion im Wesentlichen destruktiver Natur sind. Andere Evolutionsfaktoren, die im Rahmen jüngerer Hypothesen wie u. a. der „extended evolutionary synthesis“ (EES) angeführt werden, beurteilte Behe im Vergleich zur klassischen Theorie als noch weniger erklärungsstark. Darauf reagierten zahlreiche naturalistisch gesonnene Naturwissenschaftler mit zum Teil vehementer Kritik. Im Rahmen einer philosophischen Kritik wurde die Terminologie von Behe und anderen ID-Befürwortern für unangemessen erklärt, während die naturwissenschaftlichen Kritikpunkte sich auf Beispiele aus der Biologie fokussierten, die die Argumente der ID-Befürworter entkräften sollen. Auf der Ebene der Philosophie werden suggestive Argumente angeführt und von ID-Befürwortern Unmöglichkeitsbeweise gefordert. Auf der naturwissenschaftlichen Ebene wird behauptet, dass wesentliche Argumente der ID-Verfechter längst widerlegt seien und dass degenerative Evolution überbewertet würde. Es zeigt sich jedoch, dass die Einwände der Evolutionsbefürworter mangelhaft begründet sind und sich anhand einer genauen Betrachtung der Daten entkräften lassen.
Einleitung
Unser Selbstverständnis ist in einem hohen Maße davon abhängig, wie wir unsere Geschichte verstehen. Daher ist die Deutungshoheit bezüglich der Narrative über unsere Herkunft und Geschichte stets umkämpft. Das gilt auch für das große Bild der Naturgeschichte, wie verschiedene Debatten seit der Zeit Darwins zeigen. Große Bekanntheit erlangte die WilberforceHuxleyDebatte aus dem Jahr 1860 in Oxford. Der Professor für Mathematik und Bischof Samuel Wilberforce argumentierte für Schöpfung, während der Biologe Thomas Henry Huxley für Evolution eintrat. Huxley verwendete in dieser Debatte das Argument der ewig lebenden Affen, die durch Tippen auf einem unendlichen Vorrat an Papier irgendwann auch zufällig den 23. Psalm zustande brächten. Auf diese Weise wollte Huxley seine Behauptung untermauern, dass selbst komplexe Lebewesen ohne Schöpfer entstehen könnten. Da Wilberforce das Argument nicht überzeugend entkräften konnte, folgte das Publikum der Logik von Huxley. Die Inhalte der Debatte wurden weit verbreitet und dies trug dazu bei, dass die biblische Schöpfungslehre in akademischen Kreisen als überholt galt.
Im Rahmen der „Huxley Memorial Lecture“ (1986) fand eine Debatte mit vier Diskutanten statt. Edgar Andrews und Arthur E. Wilder-Smith vertraten die Schöpfungslehre, während John Maynard Smith und Richard Dawkins die Evolutionslehre befürworteten. Diskutiert wurde die These, dass „[d]ie Doktrin der Schöpfung wissenschaftlich besser begründet sei als die Theorie der Evolution“. Am Ende der Debatte erhielten Andrews und Wilder-Smith 115 von 300 Stimmen – was angesichts des durchweg naturalistisch gesonnenen Publikums eine große Überraschung war. Laut Wilder-Smith gab es anschließend jedoch keine Medienberichte über die Diskussion und die Protokolle der Veranstaltung waren aus dem Archiv der Oxford-Universität verschwunden (Wilder-Smith & Wilder-Smith 2000). Wilder-Smith kommentierte dies mit den Worten: „So vollkommen ist die heutige Zensur von jeglicher effektiven Kritik an neodarwinistischer Wissenschaft und von jeder dargebotenen echten Alternative“ (Wilder-Smith & Wilder-Smith 2000). Im Unterschied zu der Wilberforce-Huxley-Debatte wurden die Verbreitung der Debatte und die sachliche Aufarbeitung der Argumente unterdrückt.
Die Feststellung von Wilder-Smith bezog sich auf eine Zeit, als die Befürworter der Schöpfungslehre* [Sternchen: s. Glossar unten] bereits eine kleine Minderheit in Europa waren. Damals blieb die Empörung aus, als die Debatte verschwiegen wurde. Inzwischen sind Meldungen aus Europa zur Frage nach Schöpfung oder Evolution eher selten geworden. Wenn zuletzt in den Medien über die Schöpfungslehre berichtet wurde, dann ausschließlich in ausgesprochen negativem Licht. Eine sachliche Auseinandersetzung mit den Argumenten findet in der Öffentlichkeit fast gar nicht mehr statt. Anders verhält es sich dagegen in den USA. Laut letzter Gallup-Umfrage aus dem Jahr 2019 stimmten 40 % der befragten USAmerikaner der Aussage zu, dass der Mensch in der jetzigen Form von Gott erschaffen worden sei, während etwa 20 % die Darwin’sche Evolutionslehre befürworteten (Brenan 2019). Ein Ignorieren der Schöpfungslehre ist demnach nicht möglich. Öffentlich geführte Debatten um Schöpfung und Evolution sind dort daher unvermeidbar.
Zuletzt entzündete sich eine argumentative Auseinandersetzung um das jüngste Buch des Biochemikers Michael Behe mit dem Titel „Darwin Devolves“ (Behe 2019a). Unter „Devolution“ ist im Kontext des Buches eine Degeneration zu verstehen: ein zerstörerischer Vorgang, der den Verlust oder die Beschädigung von Erbinformation und den dazu gehörenden biologischen Funktionen zur Folge hat. Laut Behe, der seit über 30 Jahren an der Lehigh-Universität (Pennsylvania) als Wissenschaftler tätig ist, sind die zentralen Evolutionsfaktoren Mutation und natürliche Selektion im Wesentlichen destruktiver Natur, weswegen sie keine komplexen biologischen Funktionseinheiten hervorbringen können. Evolution geschieht demnach nicht durch Aufbau neuer genetischer Information, sondern durch deren Verlust. Der
Autor bringt in der „ersten Regel der adaptiven Evolution“ zum Ausdruck, welche Vorgänge auf der Ebene der DNA für den Fitnessgewinn von Lebewesen ursächlich sind (Behe 2019a, S. 185):
„Das Zerstören oder Beschädigen eines funktionellen codierten Elements, dessen Verlust einen Fitnessgewinn nach sich zieht.“
Es liegt nahe, dass das im Gefolge Darwins allgemein akzeptierte Bild der Naturgeschichte – eine unmerklich langsame Höherentwicklung durch Mutation und Selektion und andere Faktoren vom Bakterium bis zum Blauwal im Verlauf von vier Milliarden Jahren unter erheblicher Zunahme an genetischer Information – mit der in „Darwin Devolves“ dargelegten Sicht nicht kompatibel ist. Das gilt auch für neuere Vorstellungen sprunghafter Evolution.
Die in „Darwin Devolves“ dargelegte Sicht einer stark überwiegend destruktiven Mikroevolution ist mit der allgemein akzeptierten Höherentwicklung der Organismen nicht kompatibel.
Bereits in früheren Veröffentlichungen wie „Darwin’s Black Box“ (Behe 1996) und „The Edge of Evolution“ (Behe 2007) wurde überzeugend dargelegt, warum ungesteuerte evolutive Vorgänge als Erklärung für das Phänomen der nichtreduzierbaren Komplexität* (NK) in der Biologie unzureichend sind. In seinem zuletzt veröffentlichten Buch jedoch verschärfte der Autor seine Argumentation weiter, indem er die natürlichen Evolutionsfaktoren zur Erzeugung funktionaler Information* nicht nur für ungenügend, sondern für überwiegend zerstörerisch erklärte. Damit änderte sich auch seine Einschätzung der Reichweite der Darwin’schen Evolution. Während er in seinen früheren Werken noch davon ausging, dass intelligente
Schöpfung für Phänomene wie die Feinabstimmung der Naturkonstanten, die Entstehung biochemischer Maschinen der Zelle sowie Tierklassen erforderlich sei, zog Behe diese Grenze nunmehr auf der Ebene von Tierfamilien (Behe 2019a, S. 156). Dies stellt inhaltlich eine starke Annäherung an die biblische Schöpfungslehre bzw. Grundtypenbiologe* dar, obwohl dies nirgends ausdrücklich kommuniziert worden ist.
Aufgrund der ausgewiesenen Expertise des Autors, seiner argumentativen Kraft und der weiten Verbreitung seiner Werke konnte die Fachwelt seine Publikationen nicht ignorieren. Es ist daher nicht überraschend, dass eine breite Front an naturalistisch gesonnenen Wissenschaftlern mit zum Teil harscher Kritik auf die Veröffentlichung von „Darwin Devolves“ reagierte, worauf Behe wiederholt antwortete. Im Unterschied zu den zuvor erwähnten öffentlichen Debatten wurde diese ausschließlich in Form schriftlicher Veröffentlichungen geführt. Die Relevanz der Auseinandersetzung zeigt sich darin, dass sogar in der renommierten Zeitschrift Science eine kritische Rezension von „Darwin Devolves“ veröffentlicht wurde (Lents et al. 2019). Eine Analyse des argumentativen Austauschs bietet nicht nur eine Übersicht über die vorgetragenen Argumente und den Stand der Debatte, sondern offenbart auch einiges über die Haltung und Vorgehensweise der Befürworter der jeweiligen Sichtweise.
Kritik an Behes jüngstem Werk „Darwin Devolves“ und Erwiderung
Wenn man die Kritik an Behes Werken allgemein beschreiben wollte, so wären Begriffe wie „allumfassend“ oder „breit angelegt“ zutreffend. Auch wenn der Kern der Debatte eine naturwissenschaftliche Fragestellung ist, beschränken die Kritiker ihre Äußerungen keineswegs auf diesen Bereich.
Kritik auf sprachlicher Ebene: die verwendeten Begriffe
Ein nicht unerheblicher Teil der Auseinandersetzung ist auf der sprachlichen MetaEbene verortet: Der Kampf um die Deutungshoheit setzt bei der Wahl von Schlüsselbegriffen ein, weil sie für das Denken richtungsweisend sind. Der Biologe Nathan H. Lents von der Universität New York bezeichnete in einem seiner Blogbeiträge die Wahl des Begriffs „Devolution“ als „sinnlos“ und einen „puren Beheismus“ (Lents 2019). Er begründete das damit, dass angeblich niemand in der Wissenschaft diesen Begriff gebrauche, noch verstehe. Lents fügte dem noch hinzu, Behe würde die Natur fundamental falsch verstehen, indem er mit dem Begriff „Evolution“ eine Richtung verbinde – eine Anhäufung von Komplexität bzw. eine Entwicklung zur Vollkommenheit hin. Vielmehr sei Evolution ziellos und würde oftmals sowohl zu plumpen als auch zu eleganten Lösungen führen.
Lents Argumentation ist in mehrerer Hinsicht irreführend. Abgesehen davon, dass Mehrheitsargumente per se nicht überzeugend sind, zeigt Lents in seinem Beitrag, dass er selbst sehr wohl verstanden hat, was der Begriff „Devolution“ bedeutet. Er werde begrifflich der Evolution gegenübergestellt – ebenso wie Degeneration durch Genverlust das genaue Gegenteil von der Höherentwicklung durch Gewinn an genetischer Information und den dazugehörigen Funktionen ist. Es ist unbezweifelbar, dass Darwin die winzigen mikroevolutiven Veränderungen als konstruktiv auffasste und in der Summe als Ursache von Makroevolution, d. h. Höherentwicklung, sah – eine Vorstellung, die bis heute für das Denken der Biologen prägend ist. Behe dagegen äußerte sich nicht ausführlich zu Makroevolution, sondern beschränkte seine Darlegungen auf mikroevolutive Vorgänge, die ihm zufolge genetische Information langfristig zerstören und folglich keine Erklärung für den Ursprung und die Entstehung der Arten sein können. Insofern ist Devolution als konzeptionell zu Evolution entgegengesetzter Begriff sprachlich angemessen. Außerdem hätte eine einfache Stichwortsuche im Internet genügt, um auf wissenschaftliche Veröffentlichungen zu stoßen, die den Begriff „Devolution“ bzw. „devolvieren“ in genau der Bedeutung verwenden, wie es auch Behe tat (MacLean & Dickerson 2019; Benner 2017): Verlust von Information bzw. Funktionen.
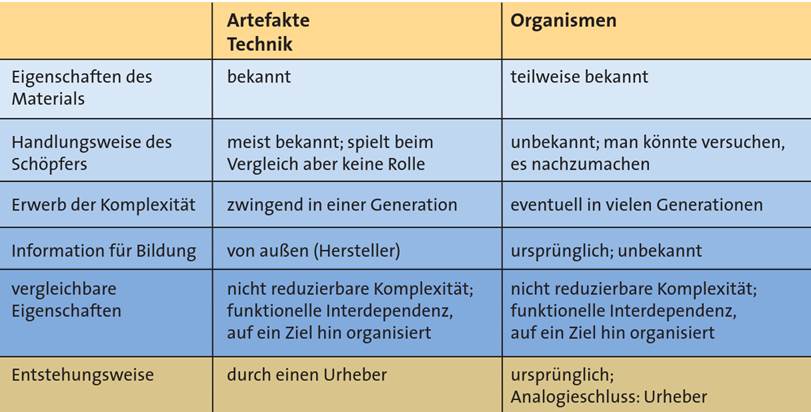
Abb. 1 Vergleich von Organismen und technischen Gegenständen zur Begründung von Analogieschüssen auf intelligente Verursachung in Lebewesen. (Aus Junker & Scherer 2013, S. 334 bearbeitet)
Kritisiert wurde auch der Gebrauch von Metaphern, die als Hinweis auf intelligente Schöpfung in der Natur aufgefasst werden können. Zwei ebenfalls an der LehighUniversität tätige Kollegen stellten in ihrer Buchrezension zu „Darwin Devolves“ den Realitätsbezug einiger gängiger Sprachbilder in Abrede (Lang & Rice 2019). Sie unterstellen Behe die Absicht, seine Leser durch effektiven Gebrauch von Metaphern von der biologischen Realität abzulenken, um für Design zu argumentieren. Dabei seien Proteine keine Maschinen und das Flagellum kein Außenbordmotor, so die Kritiker. Behe selbst antwortete auf diesen Einwand, indem er darauf verwies, dass Proteine – und erst recht Proteinkomplexe – tatsächlich Maschinen sind, die sich nur durch ihr Material von menschengemachten Maschinen unterscheiden (Behe 2019). Auch wenn sie nicht aus Stahl, Holz oder Plastik bestehen, sind sie in gleicher Weise wie technische Maschinen aus einzelnen zweckdienlichen Komponenten zusammengesetzt und verrichten mechanische Arbeit. Das beschreiben auch des Kreationismus völlig unverdächtige Buchautoren entsprechend (Steven et al. 2016):
„Insgesamt betrachten wir eine Klasse – makromolekulare Maschinen, die als Konstruktionen betrachtet werden, deren biologische Aktivitäten einen starken mechanischen Aspekt haben und die, wie makroskopische Maschinen, mechanische Arbeit verrichten.“ (Hervorhebung B. S.)
Ein grundsätzliches Ablehnen von Sprachbildern und Analogien, die intelligente Schöpfung in der Natur implizieren, würde eine unzumutbare Beschränkung des Denkens und der Sprache bedeuten.
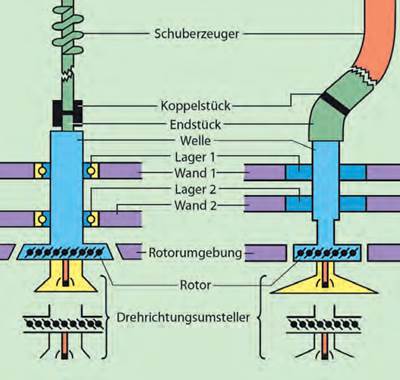
Abb. 2 Veranschaulichung der Analogie zwischen einer technischen Konstruktion und einer Entsprechung in einem Organismus. Links ist das Schema eines technischen Rotationsmotors dargestellt und rechts das Schema des Flagellums eines E. coli-Bakteriums. (Aus Junker & Scherer 2013, S. 331)
Es fällt schwer zu verstehen, weshalb Lang & Rice (2019) zu einer derart offenkundig widerlegbaren Argumentation greifen. Und auch ihre Kritik an der Analogie von Flagellum und Außenbordmotor ist in keiner Weise gerechtfertigt. Wenn Behe und andere Befürworter der intelligenten Schöpfung davon sprachen, dass ein Flagellum ein Außenbordmotor „ist“, dann im Sinne eines plausiblen Analogieschlusses, der stets aufgrund der augenfälligen Ähnlichkeit der baulichen Komponenten der beiden verglichenen Gegenstände (Rotationsachse, Ringe, Stator, Kühlung etc.) und ihrer Funktion erfolgte.
Lehnt man solche naheliegenden und treffenden Vergleiche ab, müsste konsequenterweise auf sehr viele hilfreiche Analogien verzichtet werden, selbst wenn sie sich allen Betrachtern aufdrängen. Das wäre eine unzumutbare Beschränkung der Sprache und des Denkens, die sicherlich kaum jemand konsequent einhalten könnte. Beispiele für solche Analogien in der wissenschaftlichen bzw. explizit evolutionsbiologischen Literatur sind Legion (Junker 2021). Selbst Richard Dawkins griff bei der Beschreibung des Erbgutes zu einer Sprache, die nicht anders klingt als bei ID-Befürwortern (Dawkins 1996): „Was sich im Kern jedes lebenden Dings befindet, ist nicht ein Feuer, nicht warmer Atem, nicht ein ‚Funken Leben‘. Es sind Informationen, Wörter, Anweisungen. […] Denken wir stattdessen an eine Milliarde besondere, digitale Zeichen. […] Wenn wir das Leben verstehen wollen, [müssen wir] an Informationstechniken [denken].“
Kritik auf naturwissenschaftlicher Ebene
Nichtreduzierbare Komplexität (NK). Viele der Kritiker beginnen ihre Ausführungen mit der Behauptung, dass das Argument der NK als zentrales Konzept von ID längst widerlegt sei.
Lents et al. (2019) sprechen in ihrer Rezension dem Argument seine Gültigkeit ohne jegliche Begründung ab. Stattdessen verweisen sie auf das Urteil im Prozess Kitzmiller v. Dover im Jahr 2005, der mit einer Niederlage der IDBefürworter endete – als ob ein juristisches Urteil maßgeblich für den Ausgang einer naturwissenschaftlichen Debatte sein könnte.
Aufgrund der Wichtigkeit dieses Arguments sei es hier erneut aufgegriffen. Behe selbst definiert NK wie folgt (Behe 2019a, S. 230): „Unter nichtreduzierbar komplex verstehe ich ein einzelnes System, das aus mehreren gut aufeinander abgestimmten, interagierenden Bauteilen zusammengesetzt ist, die zur Grundfunktion dergestalt beitragen, dass das Entfernen eines dieser Bauteile zum Verlust der Funktion führen würde.“
Zur Veranschaulichung des Arguments dient dabei stets die Mausefalle, die nur bei vollständigem Vorliegen und der richtigen Anordnung der Bauteile ihre Funktion als Falle erfüllt. Da bisher kein positiver Beleg für die Möglichkeit eines schrittweisen evolutiven Zustandekommens nichtreduzierbar komplexer Systeme vorliegt, ist intelligenter Input die einzig plausible Erklärung.
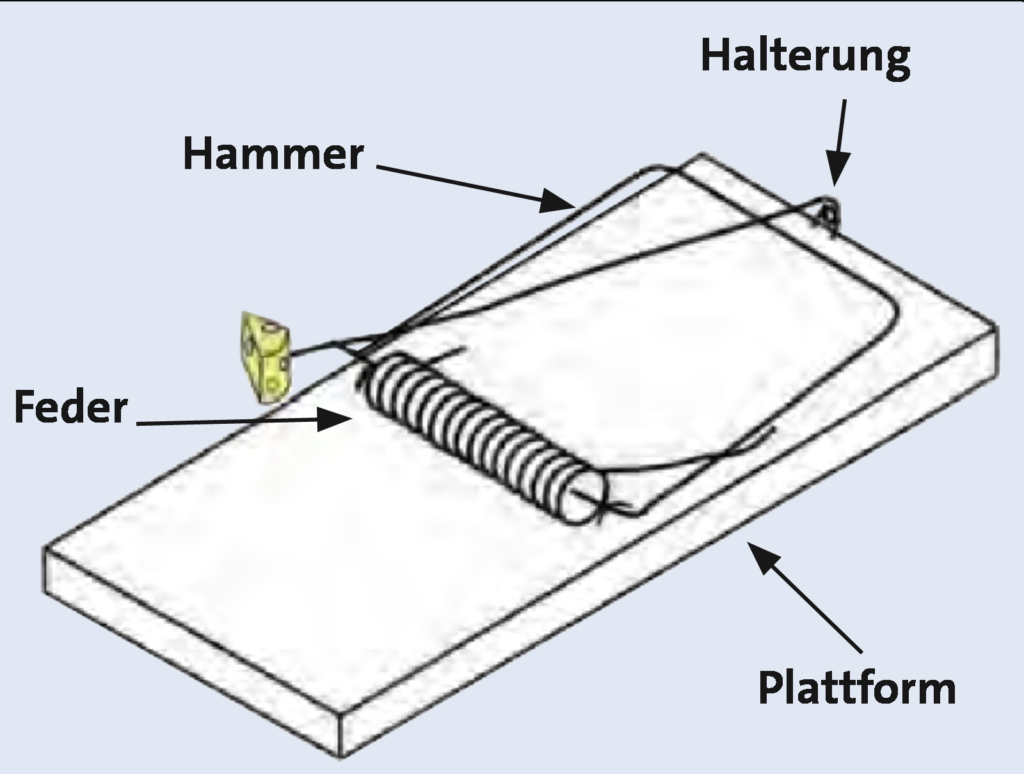
Abb. 3 Die funktionalen Bestandteile einer nichtreduzierbar komplexen Mausefalle (aus Junker 2008, Abb. 4 nach McDonald)
Nachdem schon auf vielerlei Weise versucht worden ist, das Argument der NK zu widerlegen, sahen sich auch Lang & Rice (2019) zu einem neuen Versuch veranlasst. Sie bemängeln, dass das Argument lediglich den gegenwärtigen Zustand des jeweiligen Systems in Betracht ziehe und davon ausgehe, dass die komplexe Interaktion immer bestanden habe:
„Die Tatsache, dass ein System in seiner jetzigen Form nichtreduzierbar komplex ist, ist kein Beweis dafür, dass es sich nicht durch zufällige Mutation und natürliche Selektion entwickelt hat“ (Lang & Rice 2019).
Weiter heißt es in derselben Veröffentlichung, dass multifunktionale Proteine dafür sprächen, dass unerwartete Entwicklungswege über verschiedene Funktionen möglich seien, die bei bloßer Betrachtung einer einzelnen Funktion nicht berücksichtigt worden sein könnten.
Das angeführte Argument ist keineswegs neu. Im Grunde rekapituliert es ein gedankliches Motiv, das schon Charles Darwin in Bezug auf die hypothetische Evolution des Linsenauges angeführt hat: „Jemand, der so weit gehen wird, […], sollte nicht zögern, weiter zu gehen und zuzugeben, dass eine Struktur auch so perfekt wie das Auge eines Adlers durch natürliche Selektion entstanden sein könnte, obwohl er in diesem Fall die Übergangsformen nicht kennt. Seine Vernunft sollte die Überhand über seine Vorstellungskraft gewinnen“ (Darwin 1859).
Hier handelt es sich in keiner Weise um einen tatsächlich erbrachten Beweis in Form eines beobachteten natürlichen Vorgangs, sondern um einen Appell an die Vorstellungskraft. So wie Darwin forderte, sich nicht erwiesene evolutive Wege zum Linsenauge vorzustellen, fordern Lang & Rice, dass man sich bisher nicht gezeigte, rein hypothetische Entwicklungswege von Proteinen vorstellt. Behe selbst antwortete auf den Einwand seiner Kollegen, dass das Argument der nichtreduzierbaren Komplexität nirgends fordert, dass Proteine nur eine einzige Funktion haben dürfen (Behe 2019b). Tatsächlich liegt die Beweislast hier auf Seiten der Befürworter der Evolutionslehre. Sie müssten konkret zeigen, wie ein nichtreduzierbar komplexes System durch schrittweise Mutationen und Selektion entstehen kann – unabhängig davon, ob im Ausgangszustand Multifunktionalität der einzelnen Bestandteile bereits vorliegt oder nicht. Ein solcher positiver Nachweis steht aber noch immer aus. Stattdessen fordern Befürworter der Evolutionslehre das Erbringen eines Unmöglichkeitsbeweises: ID-Befürworter sollen zeigen, dass die evolutive Entstehung nichtreduzierbar komplexer Systeme unmöglich sei. Diese Forderung ist im o. g. Zitat von Lang und Rice implizit enthalten. Das ist eine wissenschaftstheoretisch nicht berechtigte Forderung bzw. eine unzulässige Verlagerung der Beweislast.

Abb. 4 Der Weißkopfseeadler verfügt über Augen, die ihm eine äußerst präzise Sicht auf ca. 1 km Abstand ermöglichen. Darwin forderte seine Leser auf, sich Zwischenstufen der Entwicklung ausgehend von primitiven Vorstufen vorzustellen. Dieser Denktradition folgend fordern Biochemiker, sich Zwischenstufen der Entwicklung von Proteinkomplexen vorzustellen. (Pixabay)
Die Einwände gegen das Argument der NK beruhen auf Appellen, sich nichtexistente Zwischenstufen vorzustellen, oder auf fehlenden Unmöglichkeitsbeweisen einer evolutiven Entstehung von NK.
Den bisher letzten bekannten Versuch, einen positiven Beweis für die Möglichkeit einer evolutiven Entstehung von NK zu erbringen, unternahm Arthur Hunt (2007; revidierte Fassung 2019). Er führte das in Maispflanzen vorkommende mitochondriale Gen T-urf13 an, das für ein Transmembran-Protein (ein Proteinkomplex) codiert, welches als gesteuerter Ionenkanal fungiert. T-urf13 wurde bei Maispflanzen nachgewiesen, die auf Sterilität ihrer männlichen Blüten hin gezüchtet worden waren. Diese Züchtungsvariante erwies sich allerdings als sehr anfällig für den Pilz Cochliobolus heterostrophus, dessen Giftstoff bei den Maispflanzen eine Krankheit auslöst, da er durch Binden an den Ionenkanal dessen Öffnung verursacht (Turgeon & Baker 2007). Dies hat zur Folge, dass für die Funktionsweise des Mitochondriums unverzichtbare Ionengradienten abgebaut werden. Es konnte gezeigt werden, dass sowohl die Sterilität männlicher Maisblüten als auch die Anfälligkeit für die Pilzerkrankung mit der besagten Genvariante in Verbindung stehen.
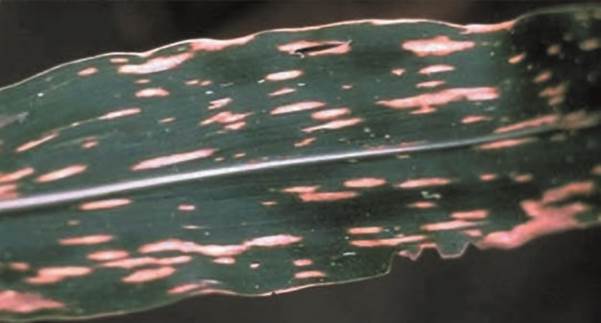
Abb. 5 Befallenes Blatt einer Maispflanze mit Cochliobolus heterostrophus. (Wikimedia: David B. Langston – http://www. forestryimages.org/browse/ detail.cfm?imgnum=5076079, CC BY 3.0)
Die Ergebnisse des Genomvergleichs zwischen der für den Pilz anfälligen Maisvariante und der normalen Pflanze wurden unter Voraussetzung von Evolution so interpretiert, dass T-urf13 durch eine Reihe von Umstrukturierungen des genetischen Materials wie Duplikationen und Rekombinationen entstanden sein soll (Hunt 2007). Euphorisch kommentierte Hunt dazu: „Mit anderen Worten: Eine nichtreduzierbar komplexe Struktur ist auf einen Schlag entstanden, ausgehend von DNA-Sequenzen, die nicht für Proteine codieren. Im Grunde ist dies ein Fall von de novo entstandener nichtreduzierbarer Komplexität“ (Hunt 2007, revidierte Fassung 2019).
Allerdings fehlen die konkreten Belege dafür, dass T-urf13 tatsächlich durch die angesprochenen genetischen Umstrukturierungen entstanden ist. Der bloße Genomvergleich ist keine hinreichende Bestätigung für den angenommenen Mechanismus. Nirgends in seiner Veröffentlichung hat Hunt den schrittweisen evolutiven Aufbau von T-urf13 im Detail beschrieben. Auch ist nirgends in seinen Ausführungen die Rede von einem Selektionsvorteil, den der Erwerb von T-urf13 nach sich ziehen könnte. Das ist angesichts des doppelten selektiven Nachteils durch Sterilität männlicher Blüten und Anfälligkeit für eine Pilzerkrankung nicht überraschend. Plausibler erscheint daher die Annahme, dass das Gen T-urf13 bereits vorher vollständig vorlag und durch eine Änderung der Genregulierung aberrant* wurde (McLatchie 2019). Dies wird dadurch bestätigt, dass das Protein Rf2, welches T-urf13 reguliert und ausgehend von der KernDNA exprimiert* wird, bereits vor einiger Zeit identifiziert werden konnte (Cui et al. 1996). Es spricht also einiges dafür, dass es sich bei diesem Beispiel um eine Genveränderung handelt, die einen Funktionsverlust nach sich zieht und damit besser durch Devolution zu beschreiben wäre (McLatchie 2019). Jedenfalls scheinen weitere Untersuchungen erforderlich zu sein, um eindeutig zeigen zu können, ob T-urf13 durch eine Reihe von Umstrukturierungen des Genoms neu zustande kommen kann. Klar ist aber auch, dass es sich hier nicht um Höherentwicklung im eigentlichen Sinne handelt, da Träger dieser Genvariante aus dem Evolutionsprozess ausscheiden.

Abb. 6 Braunbär und Eisbär. Die Tiere unterscheiden sich insbesondere durch ihre Fellfarbe und den Fettstoffwechsel. Es deutet vieles darauf hin, dass sowohl die weiße Fellfarbe als auch der an die fettreiche Kost angepasste Fettstoffwechsel der Eisbären Folgen schädigender Mutationen sind. (Pixabay)
Als Beweis für die Möglichkeit eines evolutiven Zustandekommens von NK wurde ein genetisches Merkmal in Maispflanzen angeführt, dessen Träger aus dem Evolutionsprozess ausscheiden.
Beurteilung von Mutationen. Ein weiterer Einwand, den alle Kritiker wiederholt gegen Behes Schriften vorbringen, ist eine angeblich selektive Datenauswertung, die dazu diene, die Darwin’sche Evolution als insgesamt destruktiv darzustellen. Es sei zwar richtig, geben die Befürworter der Evolutionslehre zu, dass adaptive Evolution oft von Genveränderungen herrühre, die als „Funktionsverlust-Mutationen“ beschrieben werden können. Dennoch würde Behe destruktive Veränderungen des Erbguts überbewerten und angeblich nachgewiesene evolutive Innovationsmechanismen ignorieren (Lang & Rice 2019). Besonders im Fokus standen dabei Genvarianten im Erbgut von Eisbären und die Beurteilung der Ergebnisse des Langzeitexperiments von Richard Lenski.
Eisbären sind mit Braunbären verwandt. Kreuzungen zwischen den beiden Bärenarten sind möglich und geschehen in freier Wildbahn („Capuccino-Bären“). Im Vergleich zu Braunbären weisen Eisbären ein weißes Fell auf und einen Fettstoffwechsel, der ihnen eine wesentlich fettreichere Kost ermöglicht, ohne dass es dadurch zu einer stark erhöhten Sterblichkeit durch Herzerkrankungen kommt. Liu et al. (2014) beschrieben die genetischen Veränderungen, die den genannten Anpassungen bei Eisbären zugrunde liegen. Während es hinsichtlich des Verlusts der braunen Fellfarbe klar zu sein scheint, dass eine destruktive Veränderung des LYST-Gens ursächlich ist, wurde eine längere Auseinandersetzung darüber geführt, ob das auch für den veränderten Fettstoffwechsel der Eisbären gelte. Liu et al. konnten nachweisen, dass das APOBGen, welches die Synthese des Apolipoproteins B (apoB) codiert, sich bei Eisbären und Braunbären unterscheidet. Das Protein apoB ist zuständig für das Binden von Cholesterin im Blut und dessen Aufnahme in die Zellen. Mutationen im APOB-Gen sind bei einigen Säugetieren und dem Menschen bereits als Ursache von Arterienverkalkung (Arteriosklerose) und Erkrankungen des Herzmuskels (Cardiomyopathie) ausgemacht worden. Liu et al. halten sich mit einer exakten Beurteilung der genetischen Veränderungen im APOB-Gen von Eisbären zurück, schreiben jedoch, dass „für einen großen Anteil (ca. 50 %) der Mutationen eine funktionell schädigende Wirkung vorhergesagt wurde“ – wobei die Vorhersage von der Software PolyPhen2 stammt, die Mutationen entweder als „schädigend“ oder „gutartig“, d. h. nicht schädigend, einordnen kann.
In „Darwin Devolves“ wurde die Veränderung des Erbguts der Eisbären der Terminologie des Artikels von Liu et al. folgend als destruktiv eingeordnet. Kritiker wandten dagegen ein, dass gerade die Funktion des APOB-Gens, den Cholesterin-Gehalt im Blut zu reduzieren, dagegen spräche, dass destruktive Mutationen ein schnelleres Entfernen von Cholesterin ermöglichen könnten, wie es bei Eisbären der Fall ist. Dementsprechend müsste es sich um eine konstruktive Mutation handeln, die zu einer Optimierung der Proteinfunktion beigetragen habe. Außerdem könne, so die Kritiker, die von Liu et al. verwendete Software gar nicht beurteilen, ob eine Mutation konstruktiv sei, da sie nur zwischen „schädigend“ oder „nicht schädigend“ unterscheide.
Abb. 7 Strukturformel von Cholesterin, der chemischen Verbindung, welche häufig bei Menschen und Säugetieren Herzerkrankungen verursacht und deren Gehalt im Blut durch apoB-Proteine reguliert wird.
Die Einwände der Kritiker hören sich zunächst nachvollziehbar an. Doch konnte gezeigt werden, dass das APOB-Gen nicht nur für das Entfernen, sondern auch für das Einführen des Cholesterins ins Blut zuständig ist. Das Gen codiert nämlich für die zwei isoformen* Proteine apoB-48 und apoB-100, von denen die kleinere Variante das Cholesterin aus den Zellen ins Blut befördert. Ist die Funktion des Letzteren beschädigt, wirkt sich das im Sinne einer Senkung des Cholesterinspiegels aus (Whitfield et al. 2004). Diese Fragen sind bisher nicht im Detail geklärt. Doch liegt es nahe, dass eine destruktive Veränderung des APOB-Gens die plausiblere Interpretation der Anpassung des Eisbären an seine fettreiche Kost ist. In einer Antwort auf die Einwände der Kritiker listet Luskin (2019) eine Reihe von Quellen auf, die bestätigen, dass schädliche Mutationen im APOB-Gen bei Menschen für einen niedrigen Cholesterinspiegel ursächlich sind. Ob dieser Sachverhalt auch auf Eisbären zutrifft, wird die künftige Forschung zeigen müssen. Es ist jedoch allgemein bekannt, dass Genfunktionen bei allen Säugetieren und dem Menschen in sehr vielen Fällen übereinstimmen. Auf das zuletzt genannte Argument ging Lents, einer der schärfsten Kritiker Behes, auch in seinen letzten Veröffentlichungen nicht ein (2021 und 2023).
Kontrovers diskutiert wurde auch das Langzeitexperiment von Richard Lenski (long term evolutionary experiment, LTEE). Bei dem Experiment wurde das Wachstum von E.-coliBakterien unter Laborbedingungen beobachtet, mit dem Ziel, deren Evolution nachvollziehen zu können. Inzwischen sind über 75.000 Bakteriengenerationen durchlaufen (Lenski 2023), wobei regelmäßig repräsentative Proben tiefgefroren werden, um sie für Untersuchungen von Veränderungen im Verlauf der Generationenfolge verfügbar zu haben. Behe (2019a) sieht in den Resultaten des LTEE als „überwältigend wichtige und fast komplett übersehene Lektion“ eine weitere Bestätigung der konstanten Degeneration von genetischem Material. Tatsächlich ging die große Mehrheit der Mutationen bei den E.-coli-Bakterien im LTEE mit Funktionsverlusten einher – das gaben auch Lents et al. (2019) in ihrer kritischen Rezension zu.
Das Langzeitexperiment von Lenski ist eine weitere eindrucksvolle Bestätigung der konstanten Degeneration des genetischen Materials von Lebewesen.

Abb. 8 Aufnahme der von Lenksi et al. angesetzten Bakterienkulturen für das Langzeitexperiment. (Wikipedia: Brian Baer and Neerja Hajela – http://myxo.css. msu.edu/ecoli/citrateflasksphotos.html, CC BY-SA 1.0, CC BY-SA 1.0)
Lediglich eine einzige Veränderung durch eine Reihe an Mutationen wurde von Lenski als evolutionäre Innovation eingestuft: Diese ermöglichte den E.-coli-Bakterien den Konsum von Zitrat in Gegenwart von Sauerstoff (Blount et al. 2008). Normalerweise sind E.coli-Bakterien nur in Abwesenheit von Sauerstoff dazu in der Lage, Zitrat zu konsumieren. Da Zitrat im verwendeten Nährmedium aus technischen Gründen reichlich vorlag, konnte die mutierte Variante (C+) sehr viel schneller wachsen als die anderen. Auch wenn es sich bei C+ nur um eine Reorganisation eines bereits vorhandenen Genabschnitts handelt, wurde zunächst optimistisch darüber berichtet (Blount et al. 2008; Pennisi 2013). In einem neueren Artikel hieß es jedoch, dass die vorzeitige Mortalität der C+-Variante nach ca. 50.000 Generationen auf 40 % gestiegen war (Blount et al. 2008). Dies führte bei diesen Zelllinien sogar vermehrt zu einer Art Aasfresserei – die E.-coli-Bakterien verzehrten tote Artgenossen. Es liegt nahe, dass die stark erhöhte Mortalität der C+-Variante eine Folge der Anhäufung degenerativer Mutationen war – ein weiterer offensichtlicher Fall von Devolution (Behe 2020).
Außer den angeführten Beispielen für Devolution gibt es noch viele andere. In dieser Ausgabe von Studium Integrale Journal wird die inzwischen bestätigte Degeneration von Mammuts diskutiert: Schmidtgall 2023. Darüber hinaus können noch die Sichelzell-Anämie (Behe 2019a, S. 182), die Variation der Pigmentierung des menschlichen Auges (Brilliant 2001) oder auch Verlustmutationen bei der Hefe Saccharomyces cerevisiae (Sales-Lee 2021) genannt werden. Für konstruktive Veränderungen durch die Wirkung von Evolutionsfaktoren fehlen nach wie vor die Belege.
Manche Kritiker warfen Behe überdies vor, viele andere Evolutionsfaktoren wie Plastizität* oder EvoDevo* nicht gebührend berücksichtigt zu haben. Insbesondere im Rahmen jüngerer Varianten der Evolutionslehre wie der „Extended Evolutionary Synthesis“ (EES) sind einige andere Vermutungen über Quellen für biologische Innovation vorgebracht worden. Das Ignorieren dieser Faktoren sei, so der Einwand, möglicherweise auch ein Grund für Behes Gesamteinschätzung der Evolution. Allerdings widmete Behe ganze zwei Kapitel in „Darwin Devolves“ neueren Erklärungsversuchen für die Quelle der Innovation in der Biologie und konstatierte, dass keiner dieser neuen Ansätze in der Lage ist, aufzuzeigen, wie komplexe biochemische Maschinen entstanden sein könnten. In seinen Worten: „Meiner Meinung nach gewinnt Darwin den Wettbewerb um den besten der völlig unzureichenden Mechanismen“ (Behe 2019a, S. 252).
Beurteilung der Debatte
Eine Gesamtschau des argumentativen Austauschs um das Buch „Darwin Devolves“ zeigt relativ klar, dass die daran geäußerte Kritik nicht überzeugend ist. Sie erweist sich oft als mangelhaft begründet, unsachlich oder leicht widerlegbar. Behauptungen, das Argument der nichtreduzierbaren Komplexität sei widerlegt, erweisen sich als unhaltbar oder gar irreführend. Es ist bezeichnend, dass als einziges Beispiel für die angebliche Entstehung einer nichtreduzierbar komplexen Funktionseinheit das bei Maispflanzen vorkommende Gen T-urf13 angeführt wird, welches dessen Träger aus der Evolution ausmerzt. Auch die Verweise auf angeblich konstruktive Mutationen, die zu evolutiven Innovationen führen sollen, erweisen sich bei näherer Betrachtung als degenerative Veränderungen (oder bestenfalls Reorganisationen). „Devolution“ scheint nach aktuellem Wissensstand tatsächlich die treffendere Beschreibung für die Wirkung mikroevolutiver Prozesse zu sein und ist bereits vielfach bestätigt worden. Die beständige Abwehrhaltung der Befürworter der Darwin’schen Evolution gegen diese Erkenntnis scheint jedenfalls nicht durch eine plausible Interpretation naturwissenschaftlicher Daten begründet zu sein.
Interessant ist in diesem Zusammenhang, dass Behes Argumente keineswegs neu sind. Einige Befürworter der biblischen Schöpfungslehre haben schon deutlich früher thematisiert, dass die Evolution „in die falsche Richtung geht“ (Spetner 1996) bzw. gemäß der „genetischen Entropie“ degeneriert (Sanford 2014). Dass selbst Behe diese Bücher in „Darwin Devolves“ nicht zitiert, ist sachlich nicht nachvollziehbar, denn diese Literatur dürfte ihm wohl kaum entgangen sein. Es sieht danach aus, dass selbst er jeglichen Bezug zu biblisch begründeten Überzeugungen meidet, um von der Fachwelt wahrgenommen zu werden.
Während Darwins spekulative Schlussfolgerungen bezüglich einer Höherentwicklung der Lebewesen trotz systematischer Ungereimtheiten mehrheitlich für wahr gehalten und verteidigt werden, finden wissenschaftlich solide geschriebene Bücher auch ohne Bezug zur Bibel keine breite Akzeptanz, wenn die Schlussfolgerungen der naturalistischen Weltsicht* widersprechen.
Glossar: Begriffe mit Sternchen *
aberrant: Hinsichtlich seiner Funktion abweichend. exprimieren: Übersetzen der Information von der DNA in ein Protein.
EvoDevo: Jüngere Hypothese im Rahmen der Evolutionstheorie, die sprunghafte Evolution durch Mutationen in Master-Kontrollgenen (wichtige regulatorische Gene) erklärt.
funktionale Information: Gemeint sind sinnvolle Zeichensätze, die eine Funktionseinheit codieren. So wie in einem Computer jede funktionierende Software in ganz spezifischer Weise codiert sein muss, bedarf es einer spezifischen Anordnung von Bausteinen im Erbgut, damit ein Protein codiert wird. Durch ungesteuerte natürliche Vorgänge kommt so etwas nach aktuellem Wissensstand nicht zustande.
Grundtypenbiologie: Eine Lehre, die davon ausgeht, dass es von Gott geschaffene Schöpfungseinheiten gibt, die auf der Ebene der Systematik der Lebewesen (Taxonomie) in etwa den Tierfamilien entsprechen. Innerhalb der Grundtypen kann es zur Artaufspaltung kommen, die ein Ausschöpfen vielfältig angelegter genetischer Information ist.
isoforme Proteine: verschiedene Proteine, die anhand der Information von demselben Genabschnitt synthetisiert werden. Die Unterschiede resultieren aus verschiedenen Spleißvorgängen (alternatives Spleißen). naturalistische Weltsicht: Ausschließlich auf physikalischen, d. h. natürlichen Dingen beruhendes Weltverständnis, das die Existenz übernatürlicher Dinge oder Personen prinzipiell ausschließt. nichtreduzierbare Komplexität: Ein System, das aus mehreren gut aufeinander abgestimmten, interagierenden Bauteilen zusammengesetzt ist, die zur Grundfunktion dergestalt beitragen, dass das Entfernen eines dieser Bauteile zum Verlust der Funktion führen würde. Plastizität: Die Fähigkeit bestimmter Organismen, unter unterschiedlichen ökologischen Bedingungen recht verschiedene Phänotypen (äußere Erscheinung) auszuprägen. Dies beruht auf vorhandenen Genmodulen, die bei Bedarf aktiviert werden können.
Schöpfungslehre: Hier ist die allgemeine Überzeugung gemeint, dass ein übernatürlicher Schöpfer für den Ursprung des Lebens verantwortlich ist. Eingeschlossen sind also sowohl die biblische Schöpfungslehre als auch das „Intelligent Design“.
Literatur
Behe MJ (1996) Darwin’s black box: The biochemical challenge to evolution. Free Press, New York.
Behe MJ (2007) The edge of evolution. The search for the limits of Darwinism. Free Press, New York.
Behe MJ (2019a) Darwin Devolves. The new science about DNA that challenges evolution. HarperOne, New York.
Behe MJ (2019b) A response to my Lehigh Colleagues. https://evolutionnews.org/2019/03/aresponsetomylehighcolleaguespart3/.
Behe MJ (2020) The citrate death spiral. Evolution News, https://evolutionnews.org/2020/06/citrate deathspiral/.
Benner (2017) Detecting Darwinism from Molecules in the Enceladus Plumes, Jupiter’s Moons, and Other Planetary Water Lagoons. Astrobiology, 17, 840– 851.
Blount ZD et al. (2008) Historical contingency and the evolution of a keyinnovation in an experimental population of Escherichia coli. Proc. Natl. Acad. Sci. 105 (23), 7899–7906.
Brenan M (2019) 40 % of Americans Believe in Creationism, https://news.gallup.com/poll/261680/ americansbelievecreationism.aspx.
Brilliant MH (2001) The Mouse p (pinkeyed dilution) and Human P Genes, Oculocutaneous Albinism Type 2 (OCA2), and Melanosomal pH. Pigment Cell Res. 14, 86–93.
Cui X, Wise RP & Schnable PS (1996) The rf2 Nuclear Restorer Gene of MaleSterile TCytoplasm Maize. Science 272, 1334–1336.
Darwin C (1859) On the Origin of Species. Faksimile-Ausgabe, Harvard Univ. Press, S. 188.
Dawkins R (1996) Der blinde Uhrmacher. München, 2. Auflage, S. 134.
Hunt A (2007) On the evolution of Irreducible Complexity. The Panda’s Thumb blog (revidiert am 23.01.2019).
Junker R (2008) Nichtreduzierbare Komplexität, https://www.genesisnet.info/pdfs/Irreduzible_ Komplexitaet.pdf
Junker R (2021) „Baumeisterin Natur.“ Sind teleologische Begriffe in der Biologie nur Metaphern? In: Junker R & Widenmeyer M (2021) Schöpfung ohne Schöpfer? Studium Integrale. Holzgerlingen.
Junker R & Scherer S (2013) Evolution – ein kritisches Lehrbuch. WeyelVerlag.
Lang GI & Rice AM (2019) Evolution Unscathed: Darwin Devolves argues on weak reasoning that unguided evolution is a destructive force, incapable of innovation. Evolution 73, 862–868.
Lenski RE (2023) Revisiting the Design of the Long-Term Evolution Experiment with Escherichia coli. J. Mol. Evol. 91, 241–253.
Lents NH (2019) Darwin Devolves: Devolution is not a thing. https://thehumanevolutionblog.com/2019/ 02/19/devolutionnotathing/.
Lents NH (2021) Behe, Bias, and Bears (Oh My!), https://skepticalinquirer.org/2021/03/Behebiasandbearsohmy/ und (2023) https://thehumanevolutionblog.com/2023/01/11/Behebiasandbearsohmy/.
Lents NH, Swamidass SJ & Lenski RE (2019) The end of evolution? Science 363, 590.
Liu et al. (2014) Population Genomics Reveal Recent Speciation and Rapid Evolutionary Adaptation in Polar Bears. Cell 157, 785–794.
Luskin C (2019) Polar Bear Seminar: New Evidence That Michael Behe Is Right. Evolutionnews, https://evolutionnews.org/2019/05/polarbearseminarnewevidencethatBeheisright/.
MacLean KFE, Dickerson CR (2019) Kinematic and EMG analysis of horizontal bimanual climbing in humans. Journal of Biomechanics, 92, 11–18.
McLatchie J (2019) Does Turf13 Refute Irreducible Complexity? A Response to Arthur Hunt. https:// evolutionnews.org/2019/04/doesturf13refuteirreduciblecomplexityaresponsetoarthurhunt/.
Pennisi E. (2013). The man who bottled evolution. Science 342, 790–793.
Sanford JC (2014) Genetic Entropy. Feed my sheep found inc.
Schmidtgall B (2023) Mammuts: degenerierte Elefanten? Stud. Integr. J. 30, 126127.
Spetner L (1996) Not by Chance! Shattering the Modern Theory of Evolution. Thrift Books, Chicago.
Steven AC et al. (2016) Molecular biology of assemblies and machines. Taylor & Francis Group.
Turgeon BG & Baker SE (2007) Genetic and genomic dissection of the Cochliobolus heterostrophus Tox1 locus controlling biosynthesis of the polyketide virulence factor Ttoxin. Advances in Genetics 57, 219–261.
Whitfield AJ et al. (2004) Lipid Disorders and Mutations in the APOB Gene. Clinical Chemistry, 50, 10, 1725–1732.
Wilder Smith AE & Wilder Smith B (2000) Es war ein reiches Leben. SCM Hänssler.
Zurück zu „Design-Theorie“.