Natürliche Selektion
Selektion bedeutet Auslese der Bestangepassten aus einem Überangebot von Nachkommen. Sie äußert sich in unterschiedlichem Fortpflanzungserfolg. Selektion trägt zur Variation bereits bestehender Organe und Strukturen bei, kann aber keinen Beitrag zur Entstehung neuer Organe liefern. Anpassung und Neuentstehung haben nichts miteinander zu tun.
1.0 Inhalt
Im Artikel über „Mutation“ (https://genesis-net.de/e/1-3-b/2-1/) wurden die Mutationen als Quelle für evolutionäre Neuerungen vorgestellt. Es war dort auch die Rede davon, dass schädliche Mutationen wieder ausgemerzt werden. Das Entfernen nachteiliger Veränderungen wird als Selektion bezeichnet. Die nachteiligen Mutationen werden dadurch ausgemerzt, dass ihre Träger weniger oder gar keine Nachkommen haben. Selektion bedeutet also unterschiedlichen Fortpflanzungserfolg.
In diesem Artikel werden beobachtete Beispiele für das Wirken von Selektion vorgestellt und es wird erläutert, welche Schlussfolgerungen daraus gezogen werden können.
1.1 Die Selektionstheorie
Der Grundgedanke der Selektionstheorie geht auf Charles Darwin zurück. Demnach erfolgt Selektion dadurch, dass in der Regel ein Überangebot an Nachkommen erzeugt wird, aus dem auf lange Sicht durchschnittlich nur zwei Individuen pro Elternpaar überleben. In der Regel werden das diejenigen sein, die am besten an die jeweiligen Umweltbedingungen angepasst sind. Deren Merkmale bleiben also erhalten; die anderen gehen verloren und scheiden aus der Population aus: Selektion.
Selektionsvorgänge sind in vielen Studien beobachtet worden. So wurde beispielsweise beobachtet, dass sich schon nach mehreren trockenen Jahren die Schnäbel bei Darwinfinkenarten auf den Galapagos-Inseln in Anpassung an das veränderte Spektrum des Nahrungsangebots vergrößert hatten.
Ein zweites Beispiel: Leguane der Gattung Anolis wurden auf Karibik-Inseln ausgesetzt, auf denen sie vorher nicht vorkamen. Es stellten sich nach wenigen Jahren Veränderungen in der Länge der Hinterbeine ein und zwar in Abhängigkeit von der Vegetation, auf der sich die Echsen fortbewegen. Dort, wo dünnere Äste vorherrschten, waren die Beine kürzer als dort, wo dickere Äste verbreitet waren.
1.2 Voraussetzung: polyvalenter Genpool
Als Voraussetzung für solche Veränderungen ist ein polyvalenter Genpool erforderlich, d. h. die Ausgangsgruppe, aus welcher selektiert wird, muss zum einen eine gewisse genetische Vielfalt (Polymorphismus) aufweisen. Außerdem ist ein bereits vorgegebenes Variationspotential erforderlich, das – möglicherweise durch Umweltreize ausgelöst – in wenigen Generationen solche Änderungen hervorrufen kann, wie sie beobachtet wurden.
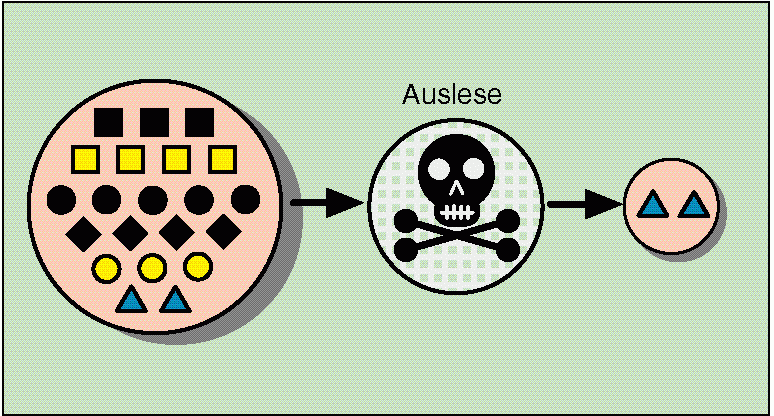
Abb. 59: Modellhafte Darstellung des Wirkens von Selektion. Aus einem reichen Genpool (links) wird nur eine (oder wenige) Variante(n) ausgelesen (Selektion). Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
Man kann solche Vorgänge wie folgt interpretieren: Aufgrund veränderter Umweltbedingungen sind aus dem Spektrum der Variabilität der Arten (Polyvalenz) andere Varianten als zuvor begünstigt. Diese können folglich besser überleben und mehr Nachkommen produzieren. Neues wird dabei nicht erzeugt, sondern aus Vorhandenem das Geeignete ausgelesen (vgl. Abb. 57, Abb. 59). Auslese kann daher nicht als Beleg für Höherentwicklung gewertet werden, denn Auslese ist ja in keiner Weise kreativ – im Gegenteil: Auslese führt dazu, dass aus dem Genpool der verschiedenen Varianten nur ein Teil in der nächsten Generation übrig bleibt. Kurz: „Auswählen“ ist nicht „erschaffen“.
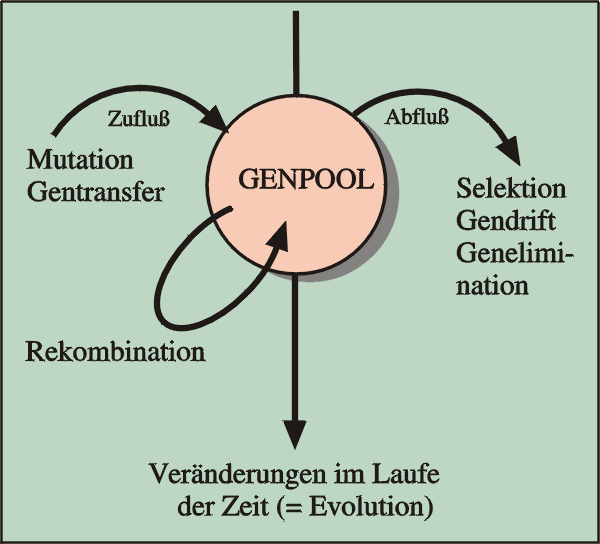
Abb. 54: Die wichtigsten Variationsfaktoren. Selektion bedeutet bildhaft gesprochen „Abfluss“ aus dem Genpool; ein Teil der vorhandenen Vielfalt wird „ausgesiebt“. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
1.3 Das berühmteste Auslese-Beispiel: Der Birkenspanner
Die Ausbreitung dunkler Formen des Birkenspanners (Abb. 60) zuungunsten heller Formen gilt als Paradebeispiel für beobachtete Evolution. Zwischen 1850 und 1920 hatten die dunklen Formen des Falters erheblich zugenommen und die hellen entsprechend abgelöst. Nach 1950 drehte sich dieser Trend wieder um. Als Ursache für die Verschiebungen der Häufigkeiten wurde die Luftverschmutzung angenommen, die zum Absterben der hellen Flechten auf den Borken der Bäume führte. Das wiederum führte dazu, dass die dort ruhenden hellen Formen des Birkenspanners nicht mehr gut getarnt waren und häufiger von Vögeln erbeutet wurden: Selektion in Aktion. Es wurde z. T. sogar die Auffassung vertreten, dass sich hier ein „prächtiges Stück Evolution“ abspielte, dass man hier „der Evolution zusehen“ durfte.

Abb. 60: Helle und dunkle Form des Birkenspanners. Es hat sich aber herausgestellt, dass sich die Falter kaum so exponiert auf Baumrinden niederlassen. Das Bild ist gestellt. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
Dieses Szenario wurde schon länger kritisiert. Der schwerwiegendste Einwand ist, dass sich nach eingehenden Feldstudien die Birkenspanner nahezu niemals auf Baumstämmen niederlassen (Abb. 60 ist gestellt). Die Ruheplätze des Birkenspanners sind bis heute nicht bekannt. Außerdem nahmen die hellen Formen wieder zu, bevor die Flechten sich wieder auf den Borken ausgebreitet hatten. Schließlich tendieren die Falter gar nicht dazu, Untergründe zu wählen, die zu ihrer Farbe passen.
Sollte es überhaupt einen Zusammenhang zwischen der Umweltverschmutzung und den Häufigkeiten der dunklen Falter geben, so ist dieser viel komplizierter als früher angenommen und bislang noch unverstanden. Vermutlich gab es zwar Selektion, doch sind die Verhältnisse im Einzelnen recht unklar.
In jedem Fall handelt es sich um einen typischen Fall von Mikroevolution. Es ist nicht dokumentiert, woher die dunklen Formen kommen. Beobachtet wurde lediglich eine Verschiebung von Häufigkeiten der Falterformen, nicht aber die Entstehung neuer Varianten. Selbst bei Annahme einer mutativen Entstehung könnte ebenfalls nur von Mikroevolution gesprochen werden, denn auch die hellen Formen besitzen den dunkelbraunen Farbstoff Melanin, welcher für die Färbung der dunklen Formen verantwortlich ist (sie haben braune Flecken oder Bänder auf ihren Flügeln), so dass die dunklen Formen ohne Strukturneubildung entstehen konnten.
1.4 Selektion und Höherentwicklung
Selektion ermöglicht eine Anpassung der Lebewesen an veränderliche Umweltbedingungen. Nach dem klassischen Neodarwinismus soll Selektion im Zusammen-
spiel mit Mutationen aber nicht nur zu verbesserter Anpassung, sondern letztlich auch zu Höherentwicklung (Entstehung neuer Strukturen) führen. Anpassungsvorgänge und Höherentwicklung stehen jedoch in keinem erkennbaren Zusammenhang. Wenn durch Selektionswirkung eine bessere Anpassung an die Umwelt erzielt wird, so ist dies (bei Wechsel der Umweltbedingungen) grundsätzlich nur mit der Veränderung eines bereits vorhandenen Organs oder Merkmals verbunden. Doch die Entstehung eines neuen Organs kann damit nicht erklärt werden, denn Selektion kann nur dort wirken, wo bereits eine Funktion ausgeübt wird, die verbessert werden kann. Umgekehrt gilt: Evolution im Sinne der Entstehung komplizierter Organe bedeutet nicht zunehmende Anpassung. Anpassung spielt sich auf einem bestimmten Organisationsniveau ab, das durch Selektion nicht verlassen wird. „Eine Amöbe, ein Wurm, ein Insekt oder nichtplazentales Säugetier ist eben-
so angepaßt wie ein Plazentalier; wären sie das nicht, so wären sie schon längst ausgestorben“ (L. v. Bertalanffy).
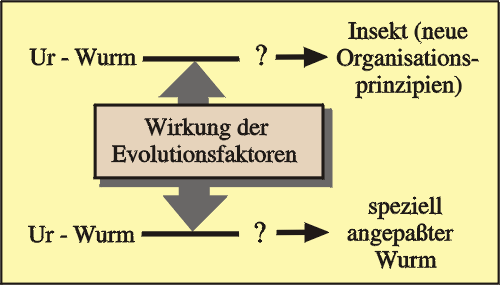
Abb. 61: Selektion macht aus einem Wurm einen besser angepassten Wurm, nicht aber ein Insekt.
Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
Ein Wurm, der wechselnden Umweltbedingungen ausgesetzt ist, wird aufgrund der Selektionswirkung zu einem „besseren“ (besser angepassten) Wurm, nicht aber zu einem Insekt. Veränderungsansätze in Richtung Insekt bedeuteten keine Anpassung als Wurm; solche Ansätze würden daher von der Selektion ausgemerzt (Abb. 61). Dieses Argument gilt auch für hypothetische gemeinsame Vorfahren von Würmern und Insekten.
1.5 Selektion: Vielfach nur behauptet
Die oben genannten Beispiele für Selektion (Darwinfinken, Leguane, Birkenspanner) gründen auf Beobachtungen im Gelände. Sie belegen offenkundig nur „Mikro- und Makroevolution“ (https://genesis-net.de/e/1-3-a/1-3/). Nun wird aber häufig behauptet, komplexe Strukturen seien ebenso durch Selektion entstanden, wie die geringfügigen Änderungen in der Schnabelgröße, Beinlänge oder Flügelfärbung bei den genannten Beispielen. Häufig wird der hypothetische Erwerb neuer Organe auch als „Anpassung“ beschrieben. Damit wird suggeriert, als könne Selektion (im Zusammenwirken mit Mutation) dafür verantwortlich gemacht werden und der Erwerb dieses Organs sei im Prinzip verstanden.
So seien beispielsweise Flügel an den Flug angepasst oder ein lange, klebrige, ausstülbare Zunge an die Ernährung durch kleine Insekten usw. Solche Ausdrucksweisen sind zwar verbreitet, aber sachlich fehl am Platz. Von Anpassung könnte man nur im Sinne einer Feinjustierung sprechen: ein bereits vorhandener Flügel kann vielleicht an verschiedene Erfordernisse beim Fliegen angepasst werden. Aber ein Flügel an sich ist aber genauso wenig eine Anpassung ans Fliegen wie eine Waschmaschine eine Anpassung an das Waschen von schmutziger Wäsche ist.
Hier ist eine angemessener Gebrauch von Begriffen wichtig: Flügel sind keine Anpassungen, sondern Einrichtungen für bestimmte Zwecke, Konstruktionen, mit denen bestimmte Aufgaben erfüllt werden können. Tatsächlich wurde die Entstehung neuer Organe durch Selektion (und andere Evolutionsfaktoren) nicht beobachtet. Selektion ist ein Variationsfaktor, jedoch kein „Kreationsfaktor“.
Autor: Reinhard Junker, 14.07.2008
Aktualisiert am 07.01.2024 (B. Scholl); © beim Autor; alter Link: 2008, https://www.genesisnet.info/schoepfung_evolution/i41242.php