Das Pfauenrad – ein Paradebeispiel für sexuelle Selektion? (Experten)

Introbild Das Prachtgefieder des männlichen Pfaus. (Bildquelle: Unsplash)
Der männliche Pfau dient als Paradebeispiel für sowohl unnötige Schönheit als auch für sexuelle Selektion (der Grundlagenartikel zu diesem Thema findet sich hier: https://genesis-net.de/sexuelle-selektion-ueberblick/). Zeugt der Pfau mit seiner enormen Schönheit nun von einem Schöpfer mit Sinn für Schönheit oder ist er das Paradebeispiel weiblicher Partnerwahl?
Zuerst zu den offensichtlichen Daten: Das Rad des männlichen Pfaus besteht aus etwa 200 einzelnen Schwanzfedern, von denen 170 einen dekorativen Augenfleck bilden; die übrigen 30 Federn werden von einem T-ähnlichen Federbusch gekrönt (Burgess 2023).
Ausgebreitet zu einem Fächer mit einem Winkel von über 180 Grad weisen jeder Augenfleck und jedes „T“ einen gleichmäßigen Abstand zueinander auf. Die kürzeren Federn sind vor den längeren angeordnet, sodass jedes „Auge“ sichtbar ist. Die Federn sind symmetrisch und geometrisch präzise angeordnet, die beiden nachfolgenden Grafiken in Abb. 4 sollen dies veranschaulichen. Gibt es für diese Muster und Geometrien plausible Erklärungen im Rahmen sexueller Selektion?
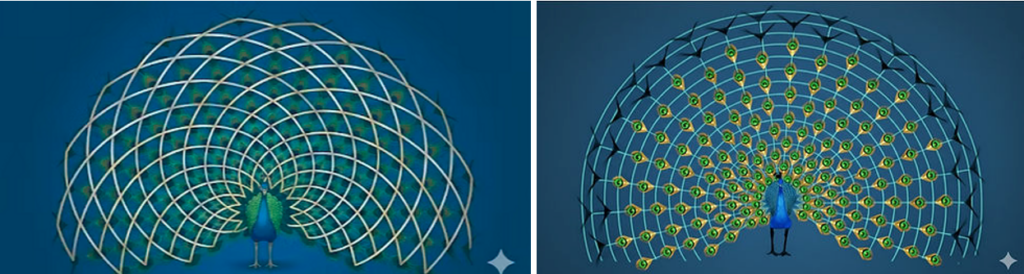
Abb. 1 Die Geometrie der Schwanzfedern. (Bilder mit KI (Gemini) erstellt)
Burgess (2023) hat sich in einem ausführlichen Artikel mit den Schwanzfedern des männlichen Pfaus und ihrer Erklärbarkeit durch sexuelle Selektion auseinandergesetzt. Er legt detailliert ihren Aufbau dar.
Augenfedern und Dünnschichtinterferenz
Im Gegensatz zu einer normalen Flugfeder verläuft der Federschaft bei einer Augenfeder nicht bis zum Ende der Feder. Er endet etwa in der Mitte, wodurch eine Teilung des Augenmusters in zwei Hälften verhindert wird. Darüber hinaus ist er im Bereich des Musters dünn, wodurch er weniger auffällt und nicht vom eigentlichen Augenmuster ablenkt, und er weist nahe des Musters eine braune Färbung auf. Letzteres ist ebenfalls für die Schönheit der Feder relevant, da ein weißer Schaft im „Auge“ zu stark auffallen würde.

Abb. 2 Schema der Schwanzfeder eines männlichen Pfaus. (Dieses Bild wurde freundlicherweise von Stuart Burgess zur Verfügung gestellt und ist hier zuerst erschienen: https://creation.com/en/articles/the-beauty-of-the-peacock-tail-and-the-problems-with-the-theory-of-sexual-selection)
Von dem Schaft zweigen die Federäste ab, die unterhalb des Musters in größeren Abständen zueinander angeordnet sind. So entsteht ein Kontrast zur Präzision der oberen Federäste, die das Augenmuster bilden. Von den Ästen zweigen wiederum die Federstrahlen ab. Am auffälligsten sind die komplexe Musterung und die leuchtenden Farben. Die Farben leuchten so schillernd aufgrund der Dünnschichtinterferenz. Dies ist ein optischer Effekt, bei dem Licht in den Federstrahlen je nach Winkel unterschiedlich gebrochen wird und so verschiedene Farben erscheinen. „Im Augenmuster erscheinen die Strahlen bronzefarben, blau, dunkelviolett und grün. Abseits der Augenregion sind die Strahlen einheitlich grün“ (Burgess 2023). Die Farben sind nur auf der Vorderseite zu sehen, wo sich die Federstrahlen befinden. Die Rückseite ist durch die Äste braun pigmentiert.
Zum Verständnis der Funktionsweise der Dünnschichtinterferenz ist zunächst Kenntnis über die detaillierte Struktur der Schwanzfeder wichtig. Wie auch gewöhnliche Flugfedern verfügen die Schmuckfedern des Pfaus über einen Federschaft, abzweigende Äste und davon wiederum abzweigende Federstrahlen. Die Besonderheit liegt in der Strahlenstruktur: Während die Strahlen bei normalen Federn schmal sind und die Äste nicht überdecken, ähneln sie beim Pfau flachen Bändern, die einander überlappen und so eine eigene Oberfläche auf der Oberseite der Federäste bilden.

Abb. 3 Vergleich der Federstrahlen von der Schwanzfeder (a) und der Flugfeder (b). (Dieses Bild wurde freundlicherweise von Stuart Burgess zur Verfügung gestellt und ist hier zuerst erschienen: https://creation.com/en/articles/the-beauty-of-the-peacock-tail-and-the-problems-with-the-theory-of-sexual-selection)
Für Dünnschichtinterferenz werden nun eine oder mehrere sehr dünne und transparente Schichten benötigt, die auf einem dunklen Hintergrund liegen. Das transparente Material muss eine zur Wellenlänge des sichtbaren Lichts ähnliche Schichtdicke aufweisen. „Sichtbare Farben haben Wellenlängen zwischen 0,4 und 0,8 µm, und Dünnfilme weisen typischerweise eine Dicke zwischen 0,3 und 1,5 µm auf“ (Burgess 2023). Darüber hinaus muss der Dünnfilm einen anderen Brechungsindex als den der Luft aufweisen, sodass das Licht beim Durchdringen des Dünnfilms verlangsamt wird. Ein bekanntes Beispiel von Dünnschichtinterferenz ist, wenn eine Öloberfläche auf nassem Untergrund blau und grün schimmert.
Beim Pfau spielt sich die Dünnschichtinterferenz in den drei die Strahlen bedeckenden Keratinschichten ab. Jeder Federstrahl hat eine Breite von etwa 60 µm und eine Dicke von 5 µm. Die Federstrahlen bestehen aus einem Schaumkern von 2 µm Dicke, der auf beiden Seiten von drei Keratinschichten umgeben ist. Die Keratinschichten sind mit einer Dicke von 0,4–0,5 µm extrem dünn.
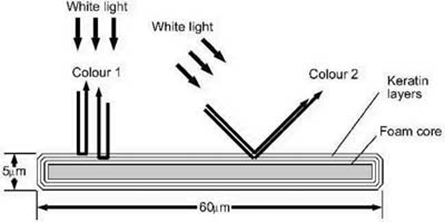
Abb. 4 Schema der Dünnschichtinterferenz. (Dieses Bild wurde freundlicherweise von Stuart Burgess zur Verfügung gestellt und ist hier zuerst erschienen: https://creation.com/en/articles/the-beauty-of-the-peacock-tail-and-the-problems-with-the-theory-of-sexual-selection)
In den Federstrahlen passiert nun Folgendes: Weißes Licht fällt auf die Keratinschichten und wird sowohl an der Vorderseite als auch der Rückseite des Dünnfilms reflektiert. Das die Keratinschicht(en) durchdringende Licht wird dabei verzögert, sodass auf seinem Rückweg einige der Farbkomponenten des weißen Lichts gegenüber den Lichtwellen, die an der Vorderseite reflektiert wurden, phasenverschoben sind.
Die Phasenverschiebung von zwei Wellenzügen derselben Farbe hat die Aufhebung der Farbe durch destruktive Interferenz zur Folge. Bei weißem Licht ist das Resultat eine Reflexionsfarbe aufgrund der übrigen Farbkomponenten des weißen Lichts.
Das einzige Pigment im Pfauenschwanz ist das Melanin in den Federästen. Durch ihre braune Farbe bieten sie einen guten dunklen Hintergrund für die Dünnschichtinterferenz. Die unterschiedlichen Farben im Augenmuster sind das Ergebnis winziger Änderungen in der Dicke der Keratinschichten, welche dafür auf etwa 0,05 µm genau sein muss.
Optimale Konstruktion der Federstrahlen
Die Federstrahlen sind damit optimal konstruiert:
- Eine optimale Dicke der Keratinschichten für die Erzeugung der leuchtenden Dünnschichtfarben.
- Die dunkelbraune Hintergrundfärbung verhindert, dass Licht durch die Feder strahlt.
- Die drei Keratinschichten fügen mehrere Lichtkomponenten hinzu und tragen so zur Brillanz der Farben bei.
- Die Strahlen sind in Längsrichtung leicht gekrümmt. Dadurch werden leicht unterschiedliche Farben vermischt, was eine Weichzeichnung der in den Keratinschichten sichtbaren Farben zur Folge hat.
Das Augenmuster
Das Augenmuster ist aufgrund seiner abgerundeten Formen besonders schön. „Die ‚Pupille‘ des Auges wird durch eine dunkelviolette Kardioide [Herzlinie] gebildet und die ‚Iris‘ durch ein blaues Ellipsoid. Diese Formen befinden sich innerhalb eines spitz zulaufenden bronzefarbenen Ellipsoids, das von einem oder zwei grünen Rändern umgeben ist“ (Burgess 2023).
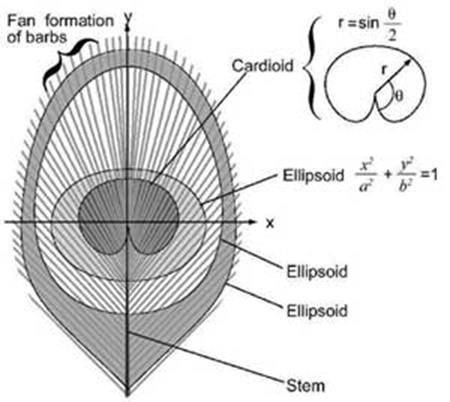
Abb. 5 Schema des Augenmusters der Schwanzfeder. (Dieses Bild wurde freundlicherweise von Stuart Burgess zur Verfügung gestellt und ist hier zuerst erschienen: https://creation.com/en/articles/the-beauty-of-the-peacock-tail-and-the-problems-with-the-theory-of-sexual-selection)
Es handelt sich beim Augenmuster um ein „digitales Muster“, da es sich aus der kombinierten Wirkung Tausender einzelner Strahlen präzise zusammensetzt. Die voneinander unabhängigen Äste müssen in ihren Positionen exakt angeordnet sein. Auch die Strahlen benachbarter Äste sind für das Augenmuster perfekt aufeinander abgestimmt. Dazu muss die Abfolge der „Farben“ bzw. interferenzwirksamen Strukturen auf jedem Ast im Erbgut vorgeschrieben sein. In der DNA müssen also Zeit- oder Positionsanweisungen vorhanden sein, die für die richtige Keratindicke am richtigen Strahl auf dem richtigen Ast codieren.

Abb. 6 Hervorhebung eines einzelnen Federstrahles mit seinen unterschiedlichen Keratindicken. (Dieses Bild wurde freundlicherweise von Stuart Burgess zur Verfügung gestellt und ist hier zuerst erschienen: https://creation.com/en/articles/the-beauty-of-the-peacock-tail-and-the-problems-with-the-theory-of-sexual-selection)
Winzige abrupte Änderungen in der Dicke der Keratinschichten der Strahlen entlang eines Astes sorgen für die klaren Grenzen zwischen den Farben. Das ist wichtig, denn graduelle Dickenänderungen würden auch fließende Farbübergänge bedeuten, wodurch das schöne Muster verschwimmen würde. Erstaunlich ist nach Burgess (2023) besonders, dass die Keratindicke entlang eines Astes nicht kontinuierlich zu- oder abnimmt, sondern in einer Richtung beide Änderungen aufweist.
Die T-Federn
Die langen T-Federn runden das Bild des aufgeschlagenen Pfauenrades perfekt ab. Die T-Federn sind zur Augenfeder invers geformt, wodurch das Innenprofil der T-Feder dem Umriss des Augenmusters folgt.
Die T-Federn bilden oftmals einen geschwungenen S-Bogen – eine sogenannte „Ogge-Kurve“ –, was ebenfalls zur Schönheit beiträgt, da sie sowohl konkav als auch konvex ist. Deshalb wird diese Form auch in der Architektur genutzt, beispielsweise beim Kielbogen. Auch diese Form ist bemerkenswerterweise wieder aus vielen einzelnen Federästen zusammengesetzt.
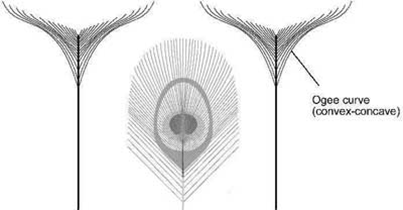
Abb. 7 Inverse Form der T-Feder zur Augenfeder und die Ogge-Kurve. (Dieses Bild wurde freundlicherweise von Stuart Burgess zur Verfügung gestellt und ist hier zuerst erschienen: https://creation.com/en/articles/the-beauty-of-the-peacock-tail-and-the-problems-with-the-theory-of-sexual-selection)
Schlussfolgerungen zur Herkunft des Pfauenrades
Die Schwanzfedern mit ihrem ausgefeilten Augenmuster spielen zweifellos eine Rolle im Balzritual. Ob Weibchen aber von ihnen „angezogen“ werden oder sie nur nüchtern als Signal zu Paarungsbereitschaft des Männchens interpretieren, ist eine andere Frage.
Offenkundig geht die Schönheit der Schwanzfedern weit über das hinaus, was für ein Signal zur Paarungsbereitschaft erforderlich wäre. Doch daraus folgt nicht objektiv zwingend eine anziehende Wirkung bei den Weibchen. Es könnte sich auch um absichtlich geschaffene Schönheit handeln, die keinen weiteren Zweck erfüllt.
Interessanterweise gibt es zu der Frage, ob die Pfauenhenne tatsächlich von den prächtigen Schwanzfedern angezogen wird, gegenteilige Studien, wie Woetzel (2017) bemerkt. Während beispielsweise Takahashi et al. (2008) in einer Studie keine Präferenz der Pfauenweibchen für längere Schwanzfedern oder eine höhere Anzahl „Augen“ nachweisen konnten, beobachteten Yorzinski et al. (2013), dass Pfauenhennen ein besonderes Augenmerk auf den unteren Teil des Federschwanzes legen.
Die wohl größte Herausforderung für eine evolutionäre Erklärung der Schwanzfedern des männlichen Pfaus liegt aber in den feinen und subtilen Details. Während Pfauen durchaus noch in der Lage sein mögen, offensichtlichere Details wie die Anzahl der Augen-Federn zu erkennen, gibt es keinen Nachweis, dass Pfauenhennen auch subtile Details wie die braune Färbung des Federschaftes am Ende registrieren können (Burgess 2023; Woetzel 2017). Da Pfauenhennen diese feinen Details also wohl auch nicht bevorzugen können, hätten sie evolutiv ohne Selektionsdruck auch nicht entstehen können – jedenfalls, solange sie nicht genetisch mit anderen Merkmalen gekoppelt sind (Pleiotropie = der Effekt, wenn ein Gen mehrere Merkmale steuert). Burgess (2023) benennt und diskutiert noch weitere Probleme.
Wenn man die hochgradige Ordnung und Präzision sowohl des Gesamtbildes (geometrische Anordnung der Federn) als auch der Details (z.B. Dünnschichtinterferenz) betrachtet, liegt es auf der Hand, dass solche feinen, abgestimmten Strukturen eher bei einem Designer zu erwarten sind als bei ungerichteten und ziellosen Naturprozessen.
Das Prachtgefieder des männlichen Pfaus kann durch sexuelle Selektion nicht plausibel erklärt werden; es ist und bleibt ein Paradebeispiel für ein Schöpfungsindiz, das als „unnötige Schönheit“ bezeichnet werden kann.
Zu grundsätzlichen Überlegungen bezüglich Sexueller Selektion: Siehe den Übersichtsartikel (Experten).
Literatur
Burgess S (2023) The beauty of the peacock tail and the problems with the theory of sexual selection. creation.com vom 05.08.2023, https://creation.com/en/articles/the-beauty-of-the-peacock-tail-and-the-problems-with-the-theory-of-sexual-selection. Erstveröffentlichung 2001 im Journal of Creation 15, 94–102.
Takahashi M et al. (2008) Peahens do not Prefer Peacocks With More Elaborate Trains. Animal Behaviour 75 (4): 1209–1219.
Woetzel D (2017) The Origin of Beauty: Contrasting the Handicap Hypothesis and Design Theory. Answers Research Journal 10, 285–292, https://answersresearchjournal.org/origin-beauty-handicap-hypothesis-design/.
Yorzinski JL et al. (2013) Through Their Eyes: Selective Attention in Peahens During Courtship. Journal of Experimental Biology 216, 3035–3046.
Dieser Artikel wurde von Janik Neuland verfasst (13.05.2026).
Hier geht es zum einfacheren Überblicksartikel zur Sexuellen Selektion und hier zum Expertenartikel.